
蜂毒肽与二元脂膜相互作用过程的单分子运动行为*
2020-06-05 10:27:40徐成林召杨恺2元冰2
物理学报 2020年10期
徐成 林召 杨恺2)† 元冰2)‡
1) (苏州大学软凝聚态物理及交叉研究中心,苏州 215006)2) (苏州大学物理科学与技术学院,苏州 215006)(2020年2月1日收到; 2020年3月31日收到修改稿)
蜂毒肽等抗菌肽可通过破坏细菌的细胞膜而直接杀灭细菌, 因此它的杀菌功能具有高效、广谱和不易产生耐药性等特点, 并被认为有望能从根本上解决目前正严重威胁人类健康的抗生素耐药性问题. 然而, 抗菌肽通过增强细胞膜的通透性来实现杀菌的分子机制至今尚不清楚. 本文结合单分子荧光追踪技术和分子动力学模拟, 从单分子运动的角度对蜂毒肽与二元脂质细胞膜的界面相互作用过程进行了研究. 结果表明, 相较于其他大多数脂分子而言、部分脂分子在膜内的扩散速率会由于蜂毒肽的吸附、聚集、插膜成孔等扰动行为发生显著降低; 而蜂毒肽倾向于作用在多组分脂膜相畴的边界处, 干扰并降低脂膜相分离的程度, 进而降低相边界对脂质分子扩散运动的限制. 本工作阐明了蜂毒肽的膜作用活性对脂分子运动行为和脂膜相行为的影响、以及三者之间的关联. 这些结果对于从单分子运动行为的角度来探索抗菌肽生物活性的分子机制和开发新型抗菌药物具有重要的指导意义.
1 引 言
细胞膜是分隔细胞内部与外界环境的界面, 更是许多重要的生物学过程发生和药物发挥作用的主要场所. 在这些生物学过程中, 抗菌肽与细胞膜的相互作用近年来广受关注. 抗菌肽是生物体免疫系统为抵抗外界细菌入侵或杀灭被感染的细胞而分泌的一类短肽, 通常包含20—60个氨基酸残基、带正电[1,2]. 与大多数传统抗生素的作用机制不同, 抗菌肽通过破坏细菌内最保守的结构之一-细胞膜, 并进而导致细菌内容物的泄漏来杀灭细菌[3],因此其杀菌功能具有高效、广谱和不易产生耐药性等特点. 因此, 面对目前正在严重威胁全球人类健康的抗生素耐药性甚至“超级细菌”问题, 抗菌肽被认为是一类有效而根本的解决途径[4,5]. 然而天然抗菌肽具有工作浓度高、细胞毒性大等缺点, 限制了其临床应用[1,4], 解决这些瓶颈问题的关键在于深入了解抗菌肽与细胞膜相互作用的分子机制.
蜂毒肽是最典型的抗菌肽种类之一, 包含26个氨基酸残基, 带6个正电荷. 根据目前普遍接受的观点[6], 蜂毒肽与细胞膜的相互作用过程大致遵循“两态”模型: 当吸附到膜表面后, 蜂毒肽会由原本的无规卷曲变为a 螺旋结构(附录A 中图A1);随后这些吸附的多肽会进一步插入膜中, 形成跨膜孔和导致跨膜物质泄漏. 蜂毒肽从表面吸附到跨膜插入的转变是其发挥生物活性的核心, 需要克服较高的能量势垒(22—37kBT)[7], 而多肽在膜表面的吸附和局部高浓度聚集以及由此对膜结构和性质造成扰动是实现这一转变的关键[7]. 实验结果显示,蜂毒肽导致膜孔形成的临界浓度约为3.0 µg/mL[8];在此临界浓度附近, 蜂毒肽在脂膜表面的吸附会导致脂膜厚度变薄约3%, 面积延展约4%[7]. Mishra等[9]利用X 射线衍射技术, 证明在多肽作用下、脂膜重组和负高斯曲率结构的出现与多肽的成孔功能密切相关; Weisshaar 等则利用荧光成像等技术证明蜂毒肽对细菌膜的成孔作用是一个反复透化、愈合、再透化的动力学过程; Huang 等[10]和Rzepiela 等[11]的模拟结果显示, 抗菌肽在膜表面的吸附及其局部浓度的增加, 会改变膜表面的压力分布, 导致磷脂尾链有序程度降低; 而我们之前的研究结果也证明, 聚集的多肽团簇能够选择性地从脂膜上叶抽离脂质分子, 造成脂膜两叶间分子成分和力学状态的不对称性[8]. 这些多肽对膜结构的扰动, 结合多肽自身的聚集和构象变化 (例如U 形弯折), 能够显著降低蜂毒肽跨膜插入的自由能势垒[12].然而, 这些相关的实验研究结果大都是从脂膜和多肽的结构变化的角度出发, 对多肽与膜相互作用过程中大量分子平均效果进行分析和对首末状态进行刻画 (例如对跨膜泄漏效果的表征和膜孔结构的分析). 此外, 大多数研究是基于单组分脂质膜模型[13], 并未涉及真实细胞膜体系内不同种类脂质分子之间的复杂相互作用对脂膜物理化学性质及脂膜与外界物质相互作用过程 (例如不同脂质分子在膜平面内的分相行为或所谓的侧向不均匀性的出现、脂筏或小窝的形成)的影响[14].
近年来, 单分子追踪及分析技术的发展使得从物理学分子运动规律研究的角度来了解生物过程的内在机制成为可能[15−17]. 本文构建了二元脂质细胞膜模型, 结合单分子荧光追踪技术和分子动力学模拟方法, 研究了不同的多肽浓度下、或多肽与脂质膜相互作用过程的不同阶段中, 蜂毒肽对脂质分子运动行为的影响. 我们的结果从单分子运动学的角度揭示了脂膜侧向不均匀性对两者相互作用的影响, 以及蜂毒肽增大细胞膜通透性和跨膜成孔的生物活性与细胞膜内的脂分子运动、以及脂膜相行为之间的重要关联.
2 实验方法与模型
2.1 材 料
磷脂分子1, 2-dioleoyl-sn-glycero-3-phosphocholine (DOPC), 1, 2-dipalmitoyl-sn-glycero-3-phosphocholine (DPPC), 1, 2-dipalmitoyl-snglycero-3-phosphoethanolamine-N-(lissamine rhodamine B sulfonyl) (Rh-PE) (分子结构式见附录A 中图A2) 购于AvantiPolarLipids. 蜂毒肽和钙黄绿素购于SigmaAldrich.
2.2 样品制备
二元脂膜以传统囊泡融合法制备.将DOPC 和DPPC 以1∶1 摩尔比混合, 加入0.1%摩尔比例的罗丹明标记的脂质Rh-PE 后在氯仿中充分溶解 (2.0 mg/mL), 利用氮气吹干并真空干燥2 h 确保氯仿完全挥发. 加入PBS 溶液后在200W 功率下超声45 min 令其重新分散为囊泡(0.1 mg/mL), 保持50 ℃(DPPC 的相变温度以上)、利用100 nm 孔径的聚碳酸酯膜挤压21次,使其成为尺寸均匀的小囊泡分散液, DLS 测定其尺寸为110 ± 15 nm. 将小囊泡分散液转移至基底表面亲水处理的玻璃仓中, 50 ℃下孵育2h 以上,最后使用1 mL PBS 溶液分三次缓慢冲洗, 即获得固体基底支撑的二元脂膜. 利用蠕动泵加入一定量蜂毒肽溶液 (浓度如正文所述)、孵育30 min 后在荧光显微镜下原位观察. 需要注意的是, 在实验中通过预掺杂少量以罗丹明化学标记的脂质RhBPE 分子来自组装制备脂膜、继而加入多肽与之发生相互作用, 这种方法可以在一定程度上避免外加染料特异性吸附的过程对脂分子运动行为的影响、以及染料分子对多肽与脂膜之间相互作用的干扰.
2.3 实验样品表征
样品的观察采用装有全内反射 (TIRF, 100 ×)镜头的荧光显微镜(IX71; Olympus)进行, 使用EMCCD(Andor DU-897U)采集图片数据, 采集速率30 fps. 每个实验体系皆进行至少3次独立重复实验, 每次实验在不同的样品或同一样品的不同区域共计采集6—8 组视频数据. 视频采集后, 采用Image J 软件MOSAIC 插件对所记录的每一个荧光标记脂质的轨迹进行分析, 从中选取有效轨迹(连续且持续时长大于3 s), 然后根据下列式子分别计算其时间平均均方位移 (time-averaged MSD,和相应的扩散系数:

2.4 模拟方法
分子动力学模拟采用粗粒化分子动力学方法,并通过Gromacs 5.1.2 运行. 脂质、水和多肽都采用MARTINI 粗粒化力场进行描述. 模拟按照Verlet 算法执行,时间步长为20 fs. 所有模拟系统都采用NPT 系综, 系统的压强设定为1 bar(1 bar=105Pa), 采用Parrinello-Rahman 方法控制; 温度采用velocity-rescale 的方法维持在300K.粗粒化粒子之间的Lennard-Jones 势在0—1.1 nm之间衰减为0. 静电相互作用计算使用reactionfield 方法, 介电常数为15. 在系统的三个方向都使用周期性边界条件.
2.5 模拟模型
根据MARTINI 力场的特点, 为了使脂膜具有更好相分离能力, 我们在模拟中采用饱和脂质DPPC、不饱和脂质dilinoleoylphosphatidylcholine(DUPC) 和胆固醇 (比例为2:2:1) 来构造脂膜.Helger 和Siewert[18]的模拟表明, 这个体系能够在有限的模拟时间尺度内实现不同脂质之间的相分离现象, 且所形成的相畴结构等特征与DPPC/DOPC 体系类似. 蜂毒肽的晶体结构可从protein data bank 网站获得 (PDB 号: 2MLT), 其粗粒化模型可进一步通过Martinize 脚本构造得到. 模拟系统中包含有12个蜂毒肽分子. 为了与实验保持一致, 我们在模拟中使用脂质相分离已经达到稳定状态的脂膜来构建初始构型, 并将蜂毒肽分子均匀放置在脂膜表面.为了考察系统在较长时间内的相互作用情况, 每一个系统都模拟了10 µs 以上.
2.6 数据处理
分别对三组实验体系下的所有有效分子轨迹进行时间平均均方位移分析, 对其扩散系数的频数分布进行多峰拟合; 利用秩和检验法确定实验结果的置信度, 利用方差检验确定小比例部分数据的统计意义; 数据显示为平均值 ± 标准差的形式 (具体的数据处理细节参见附录B).
3 结果与讨论
3.1 蜂毒肽作用导致的脂质分子运动的多样性
脂分子的流动性是细胞膜的重要特性, 受单个脂分子所处的局部膜环境和膜周围介质影响, 与细胞膜的结构、性质及行为密切相关 (例如脂膜相形为、脂筏的形成、小窝的出现、配体受体识别、细胞膜的形变及细胞迁移等)[19,20,21].对于蜂毒肽与膜相互作用体系, 脂质分子运动的变化可以直接反映多肽作用的影响, 脂分子的运动行为反过来也可能影响多肽的活性和功能. 另一方面, 浓度是影响蜂毒肽与脂膜相互作用的重要因素, 也是调控多肽与脂膜作用状态的关键. 在较低浓度下, 蜂毒肽将吸附于细胞膜的表面 (可能伴随多肽的寡聚和浅插膜行为); 当多肽浓度超过某一临界值, 多肽插膜和形成跨膜孔, 导致膜通透性增大[22,23]. 我们基于巨囊泡泄漏动力学的实验结果显示, 在3.0 µg/mL 的临界浓度之上, 蜂毒肽的加入会导致染料分子(例如钙黄绿素) 发生跨膜泄漏, 同时巨囊泡保持其连续和完整性, 意味着膜通透性增大和跨膜孔的形成;而在此浓度之下, 没有泄漏现象发生(附录A 中图A3); 当浓度高于8.0 µg/mL时, 蜂毒肽的加入会导致部分巨囊泡发生破裂. 因此在单分子运动追踪实验中, 我们选择了介于临界成孔泄漏和破膜浓度之间的5.0 µg/mL 和远低于临界成孔浓度的0.5 µg/mL 来开展实验: 按照相关文献和我们前期的研究结果[6−8], 在5.0 µg/mL 浓度下, 蜂毒肽分子应处于跨膜插入、形成膜孔的状态. 而在0.5 µg/mL浓度下, 多肽应处于脂膜表面的吸附状态.此时, 多肽会在膜表面发生肽链的聚集、构象变化, 甚至浅插入等现象, 对膜结构产生扰动[5−8]. 因此, 这一状态也等同于多肽在跨膜成孔之前的中间时间过程.总的来说, 在这两个多肽浓度下脂质分子运动的变化可以分别反映出多肽在膜吸附和导致膜通透性增大的剧烈扰动 (例如跨膜成孔) 两个作用状态下与脂膜的相互作用情况和特点.
我们利用传统囊泡融合法制备了玻璃基底搭载的DOPC/DPPC 二元单层脂膜, 以0.1%摩尔比例的Rh-PE 进行荧光标记, 荧光淬灭后恢复(FRAP) 实验验证了脂膜具有良好的流动性. 由于DOPC 与DPPC 在分子尾链结构上的差异, 此时实验中的二元单层脂膜应处于相分离状态[24].相对于常用的单组分脂膜而言, 二元相分离脂膜更有助于理解真实细胞膜体系与外界物质的相互作用过程. 之后, 我们将脂膜与0, 0.5 或5.0 µg/mL浓度的蜂毒肽溶液共同孵育30 min 后置于TIRF 显微镜下, 原位追踪单个脂分子的扩散运动轨迹. 图1(a) 是0.5 µg/mL 多肽浓度下脂膜的显微镜照片, 部分脂分子的运动轨迹典型性地显示于图1 (b)中. 由图1 (b)可以看出, 脂质分子的运动呈现出高度不均匀性. 相同时间间隔内 (例如图中轨迹时长皆为 3 s), 有些脂质运动速率快, 能够移动较大的范围; 而有些脂质的运动则仅局限于很小的范围之内, 甚至很难分辨出其位置的变化 (蓝色箭头标注). 为了对其进行定量统计, 我们计算了每个脂质分子运动轨迹对应的时间平均均方位移(MSD; 典型曲线显示于图1(c)中). 与运动轨迹分布情况相似, MSD 的分布也呈现出高度的不均匀性.
为了了解不同浓度下的蜂毒肽对细胞膜内脂分子运动行为的影响, 我们分别构建了原始脂膜以及暴露于0.5 和5.0 µg/mL 浓度蜂毒肽的脂膜体系, 并分别计算了三种体系中单个脂分子运动扩散系数(DL)的数值及其概率密度分布(PDF). 如图2所示, 对于原始脂膜, 脂质的扩散系数呈现双峰分布, 其峰位分别位于0.008 µm2/s (图2 (a) 中红色峰)和1.778 µm2/s (蓝色峰). 这说明体系中同时存在大量运动较慢和运动较快的脂分子, 这一现象与之前报道的DPPC/DOPC 二元脂膜在该摩尔比例下呈现的相分离状态是一致的(室温下DPPC应处于运动缓慢的凝胶相 (gel phase), DOPC 应处于运动较快的流体相(liquid phase))[25]. 在参照实验中我们构建了纯DOPC 脂膜体系, 其中的脂分子运动基本呈现2.780 µm2/s 位置的单峰分布(附录A 中图A4). 并且, 在该系统中荧光脂分子Rh-PE 同时存在快慢两种运动状态, 说明其分布于不同相畴区域, 这为我们研究脂膜相行为与多肽作用活性的关联提供了可能.

图1 蜂毒肽作用下脂质分子在脂膜上的运动情况 (a) 荧光显微镜照片, 部分脂分子的运动轨迹以彩色线标注; (b) 典型的脂质分子运动轨迹, 采集时间皆为3s, 部分移动范围较小的轨迹以蓝色箭头标注; (c) 典型的脂质分子时间平均均方位移分布. 蜂毒肽浓度为0.5 µg/mLFig. 1. Diffusion of lipids on a DOPC/DPPC membrane under the action of melittin at 0.5 µg/mL: (a) Microscopy image with some trajectories marked in colors; (b) typical lipid trajectories in 3 s; Some of the immobile ones are marked with blue arrows; (c) representative time-averaged MSD of lipids.
当体系中加入浓度为0.5 µg/mL 的蜂毒肽后,脂质扩散系数分布立即发生了变化. 原本的两个峰的峰位皆发生轻微左移(例如从1.778 µm2/s 降至1.660 µm2/s), 并且, 在两峰的中间出现了一个新的分布峰. 从图2(b) 中的面积比例变化可以推断,该峰的出现是由于部分快速运动的脂分子的运动速率变缓导致的(由1.778 µm2/s 降低至0.129µm2/s).根据我们之前的实验及模拟结果, 在此浓度下, 大部分蜂毒肽应呈现脂膜表面吸附的状态, 偶尔伴有瞬时孔的产生[26]. 该结果表明多肽在膜表面的吸附会限制部分脂分子的运动行为, 显著降低其扩散系数; 尤其是, 低速率新峰的出现暗示着其对脂膜相行为的影响. 继而, 在浓度为5.0 µg/mL 蜂毒肽的作用下, 脂质分子的运动发生了更为显著的变化, 三个峰皆出现明显的左移, 体系内所有分子的平均扩散系数由1.316 µm2/s 降低至0.661 µm2/s.同时, 新峰的面积比例增加(由5.49%增大到8.02%). 考虑到在此浓度下多肽已经能够形成大量跨膜孔, 该现象表明多肽导致的膜通透性增大和跨膜孔的形成会显著限制脂分子的扩散行为, 以及对脂膜相行为带来更强烈的扰动.

图2 不同浓度蜂毒肽作用下脂分子扩散系数的分布 (a) 脂质扩散系数分布柱状图及拟合曲线; (b) 相应的拟合峰中心位置和在 (a) 图中所占面积比例, 显示为拟合曲线面积比例(± 与原始柱状图面积比例之间的误差). 三个体系对应的实验样本数分别为657 (原始膜体系), 427 (蜂毒肽浓度0.5 µg/mL)和507 (蜂毒肽浓度5.0 µg/mL)Fig. 2. PDF distribution of lipid diffusion coefficients (DL) of membrane with different melittin concentrations: (a) Histograms andfittings of the PDF; (b) corresponding peak locations and area proportionsin (a). Sample numbers are 657 (pristine membrane), 427(with Mel at 0.5 µg/mL) and 507 (with Mel at 5.0 µg/mL), respectively.
3.2 蜂毒肽导致的脂质分子运动的复杂性
脂质分子在膜内的运动非常复杂. 由于所处局部膜环境不同, 不同的脂质分子侧向扩散运动能力也有所不同.我们基于小波变换方法分别分析了单个分子的运动轨迹 (附录A 中图A5—图A7 及相应方法阐述), 以获得单个脂质分子运动 (或MSD)随时间变化的情况. 结果显示, 脂质分子在膜内的运动大致分为三种模式: 某些脂质分子在整个观察时间段内持续受到移动限制, 运动速率缓慢.这表明此类脂质分子运动强烈受限或者受到外界物质的阻碍; 与之相对, 有些脂质分子在观察时间段内所受到的限制或阻碍很小, 能够保持快速运动; 此外, 也有脂质分子的运动模式会随着时间发生变化, 快、慢运动间隔发生, 说明这些脂质分子在不同的时间阶段或位置所受到的限制程度不同.相应地, 我们将脂质分子的这三种运动模式分别称为Slow 型、Fast 型和Mixed 型 (更多细节参见附录B). 总的来说, 这些脂质分子的运动模式或模式的变化都是由其所处局部环境的变化导致的, 是脂质分子与脂质分子、溶剂或外界其他物质(如多肽等) 之间复杂相互作用的直接体现.
实验发现 (图3), 对于原始脂膜体系, 约69%的脂质分子呈Mixed 型运动模式, 这说明大部分脂分子呈现快、慢交替的运动方式, 这一现象体现了脂分子运动过程中所遭受到的环境不均匀性 (例如跨越相边界). 然而, 随着蜂毒肽的加入和浓度的提高, Slow 型的脂质比例逐步增加. 这说明蜂毒肽在膜表面的吸附会限制部分脂分子的运动.而高浓度多肽对膜的剧烈扰动会增强这一限制效果, 使得更多的脂分子体现出运动受限的特性.这与前面的结果是一致的. 事实上, 与原始脂膜相比, 蜂毒肽的加入导致了Mixed 型脂分子所占比例大幅下降(由69%降至36%), 降低的该部分脂分子分别呈现了Slow 和Fast 型运动模式, 这暗示着蜂毒肽的吸附和插膜等作用倾向于发生在Mixed 型运动的脂分子区域, 降低了该部分脂分子运动过程中所遭受到的环境不均匀性, 这应是多肽对脂膜相分离行为的干扰导致的.
3.3 蜂毒肽对脂膜相边界的干扰

图3 脂质分子的三类运动模式及其在不同体系中的比例分布. 右图为典型的三类运动模式的分子轨迹, 蓝色和红色分别对应缓慢和快速运动阶段Fig. 3. Three different types of lipid diffusion modes and their PDFs in three conditions of membrane without or with melittin exposure. Representative trajectories are shown on the right.

图4 蜂毒肽对脂膜相分离行为的干扰 (a) 蜂毒肽与相分离脂膜相互作用过程的分子动力学模拟截图.上图是俯视图, 下图是侧视图. 红色对应蜂毒肽, 蓝色代表DPPC 脂分子头基, 绿色为DUPC 脂分子头基. 侧视图中脂质尾链未显示; (b) 吸附或插膜状态下多肽与周围脂质分子的相互作用细节. 颜色同 (a). 此外, 多肽周围的部分脂分子尾链以黄色棒表示, 远离多肽的脂质分子仅显示头部 (灰色); (c)相边界长度(L) 在蜂毒肽加入前后随时间的变化. 图(a) 中的时间点对应标记于图(c) 中. 插图为多肽作用下单个脂分子扩散MSD 的等值分布图, 其中以红色圆圈标出多肽位置 P/L = 12/512Fig. 4. Interaction between melittin and a phase-separated bilayer: (a) Snapshots showing the melittin-inducedporeformation process. Top: top view, bottom: side view. Red: melittin, blue: DPPC headgroup, green: DUPC headgroup. For clarity, lipid tails are not shown; (b) interaction details between Mel (red) and the surrounding lipids. Color codes are the same as in (a), with tails of the surrounding lipids in yellow, and heads of lipids away from the peptides in grey; (c) time evolution of phase boundary length (L) before and after the addition of melittin at P/L = 12/512. The time points of (a) are marked correspondingly in (c).
为了进一步验证我们的推断, 我们采用分子动力学模拟方法追踪了蜂毒肽与多组分脂质膜相互作用和成孔的过程. 如图4 所示, 原始脂膜呈现清晰的两相分离状态; 蜂毒肽的加入(多肽与脂分子之间的摩尔比为P/L=12/512) 会导致膜孔的形成.多肽与脂膜的相互作用具有两个显著的特点.其一, 多肽在膜表面会逐步聚集形成团簇, 并且, 无论多肽是处于表面吸附还是跨膜插入状态, 它都会对其周围的脂质产生强烈的吸引作用 (图4 (b)),从而显著限制吸附区域脂质的运动, 使脂分子的运动变缓(附录A 中图A8). 这与实验中观察到的脂质扩散系数下降以及快速运动脂分子比例降低的结果(图2) 是一致的. 其次, 模拟结果显示, 多肽对膜的作用显著倾向于发生在脂膜两相边界附近区域, 这可能是由于相边界区域脂膜呈现相对较多的结构缺陷供多肽实现插膜成孔功能.基于单个脂分子短时均方位移变化, 我们定量分析了由于多肽作用导致的脂膜不同区域的脂分子扩散运动变化,如图4(c)插图所示, 由图可以发现, 多肽在相边界区域的作用能起到模糊相分离的效果, 并对多肽吸附区域及周边脂分子的运动行为产生了明显的扰动.为了分析蜂毒肽对脂膜相分离行为的影响, 我们定量统计了整个相互作用过程中相边界的长度值(L) 随时间的变化, 结果显示, 在蜂毒肽的吸附和成孔过程中, 脂膜两相的边界长度出现波动并显著增大(图4 (c)). 由于在加入多肽之前脂质体系的相分离已经达到稳定状态, 因此, 此时相边界长度的变化清楚地表明了蜂毒肽对脂膜相行为的干扰和破坏性, 令两相分离程度降低.相分离行为被模糊化的现象降低了该区域脂分子环境的不均匀性, 由此降低了相边界对附近区域脂质分子运动的限制, 改变了脂质分子的运动行为规律 (例如从Mixed 模式转变为Fast 模式或Slow 模式). 这些模拟结果很好地解释了实验中观察到的由于蜂毒肽所导致的脂质分子运动行为的变化趋势(如脂质的扩散系数新的分布峰出现和Fast/Slow 模式脂质的增加等), 并证明了蜂毒肽生物活性与脂分子运动模式以及细胞膜相行为之间的关联.
4 结 论
在本工作中, 我们结合单分子荧光追踪技术和计算机分子动力学模拟方法, 从单分子运动行为分析的角度, 研究了蜂毒肽与二元脂膜的界面相互作用动力学过程, 尤其是该过程中, 处于不同作用状态的多肽(吸附于膜表面的阶段以及导致膜通透性增大和跨膜成孔的阶段) 对脂分子在膜内运动行为的影响, 以及二者之间的关联. 实验及模拟结果显示, 脂分子在二元脂膜内的运动呈现快、慢两种状态; 蜂毒肽的表面吸附会显著降低部分快速运动的脂质分子的运动速率, 而多肽对膜的进一步扰动和成孔会进一步加剧这一影响; 同时, 会令所有脂分子的运动速率发生不同程度的减慢. 并且, 蜂毒肽倾向作用于多元脂膜相分离的边界区域, 扰动并降低脂膜相分离的程度, 进而降低该区域单个脂分子运动过程中所遭受到的环境不均匀性, 降低相边界对脂质分子运动的限制. 本文从描述单分子运动行为的角度对蜂毒肽跨膜成孔实现杀菌功能的过程进行了研究, 从一个新的角度来阐述两者的相互作用分子机制. 此外, 对于今后的实验, 一个重点可以放在双色单分子成像实验上, 通过同时对多肽和脂质进行荧光标记来实现对不同组分分子的运动情况的追踪和刻画, 这将有助于全面准确地理解多肽作用和脂质运动之间的关联. 总的来说, 本工作表明抗菌肽的膜活性与脂膜的相行为密切相关,证明了细胞膜的相畴边界是抗菌肽发挥生物活性的重要作用点. 哺乳动物细胞与细菌在膜组分和结构上有明显区别, 相应的相畴结构和相边界组分也有所不同. 因此, 本文的结果表明针对这些差异来调节蜂毒肽与不同细胞膜相畴的作用方式可能是实现抗菌肽靶向选择性的有效途径之一. 这对开发新型抗菌药物具有一定的参考意义.
附录A 利用小波变换处理脂质分子运动轨迹
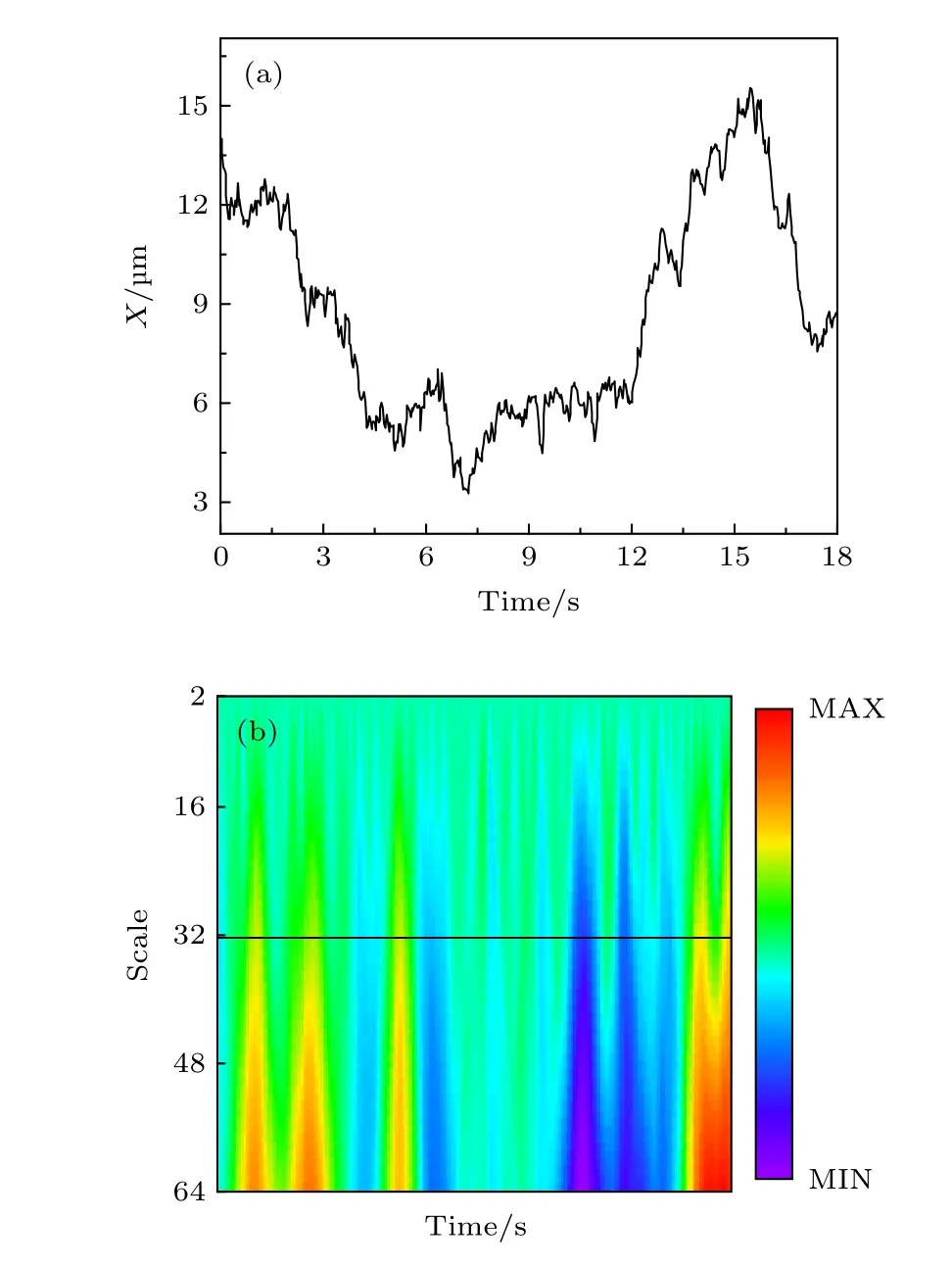
小波变换可以将一条轨迹中不同的运动模式区别开来.以图A5 (a) 所示的轨迹 (X坐标随时间的变化) 为例. 这条轨迹可以基于Haar 小波分别做第2—64 阶的变换, 其中每一阶的变换系数由下式计算得到:

图A5 (b) 显示了这条轨迹相应的小波变换系数. 对于不同阶的变换, 系数所包含的信息也有所不同, 图A6 (a)—图A6(c)分别是第2 阶、32 阶和64 阶对应的系数. 因此选择适当的系数有助于区分轨迹中所包含的不同信息. 这里, 根据相对的变化程度, 我们选择第32 阶系数作为研究对象, 通过计算阈值δ来区分轨迹中不同的运动模式. 阈值δ的计算可通过低阶变换系数 (如第2 阶) 计算得到:

图A6 (d) 显示了此轨迹经小波变换得到的第32 阶系数以及相应的阈值红线 (±δ). 红线所包含的阴影部分是轨迹中运动较慢的部分, 而在阈值以外的部分即可认为是轨迹中运动变化较快的部分. 图A7 (a)显示了经过区分后的轨迹, 红色和蓝色分别表示了轨迹中运动较快和较慢的部分.为了验证此方法的效果, 我们分别计算了轨迹中红色和蓝色部分所对应的均方位移. 如图A6 (b)所示, 两个部分的扩散方式的确有明显的区别.

图A1 蜂毒肽的 (a) 氨基酸序列和 (b) 吸附在膜表面后的a 螺旋结构Fig. A1. The sequence and a-helical structure of melittin.
图A2 DOPC, DPPC, Rh-PE 分子的结构式Fig. A2. A2. Molecular structure of DOPC, DPPC and Rh-PE.
脂分子运动模式的定量区分: Slow 型、Fast 型和Mixed 型.
我们对每一条脂质分子的轨迹进行了小波变换处理,其中红色部分即为fast 部分, 蓝色部分为slow部分, 相应地, 我们计算了这两部分轨迹的均方位移, 从中发现红色部分的斜率均大于2 (长距离扩散), 而蓝色部分的斜率均小于0.1 (短距离徘徊).
附录B 数据处理
1 数据采集及样本数
我们的实验包含三个体系, 即原始膜、加入0.5 µg/mL浓度蜂毒肽、加入5.0 µg/mL 浓度蜂毒肽体系; 每个体系分别进行了至少三次独立重复实验, 每次实验在不同的样品或同一样品的不同区域采集共计6—8个视频片段. 一般来说, 视频视野中荧光分子信号总数约为200—500个, 其中经筛选后得到的有效轨迹约占总数的25%左右 (筛选标准为轨迹必须连续且持续时长大于3 s). 每一个体系所采集到的有效轨迹的数量 (即PDF 的实验样本数) 如表B1所列.

图A3 包裹钙黄绿素的巨单层囊泡在浓度为 (a) 0.5 和 (b) 5.0 µg/mL 蜂毒肽溶液中的荧光共聚焦显微照片, 包括绿色 (钙黄绿素) 通道、红色 (脂分子) 通道和复合通道. 右图为对应的蜂毒肽作用方式的卡通图, 包括膜表面吸附和跨膜插入Fig. A3. Confocal images of calcein-encapsulated GUVs exposed to melittin at 0.5 or 5.0 µg/mL. The images were taken in the green (calcein), red (lipid) and overlaid channels. Cartoons on the right refer to the corresponding action states of peptides, including surface adsorption and transmembrane insertion.

图A4 纯DOPC 脂膜中的脂分子扩散系数分布. 实验数据来自3次独立重复实验, 样本数为404, 拟合曲线的峰位位置2.780 µm2/sFig. A4. PDF of lipids in a pure DOPC bilayer. The data were obtained from three times of independently repeated tests, with a sample number of 404. Peak of the fitted curve locates at 2.780 µm2/s.
2 实验置信度分析
我们继而使用秩和检验(Gibbons J D, Chakraborti S 2011Nonparametric Statistical Inference(5th Ed.) (Boca Raton: Chapman & Hall/CRC Press); Hollander M,Wolfe D A 1999Nonparametric Statistical Methods(Hoboken: John Wiley & Sons, Inc.)) 法对三次独立重复实验的结果进行了分析. 选择秩和检验的原因是它不依赖于总体分布的具体形式, 而t检验等一般要求数据符合正态分布.
图A5 小波变换法处理脂分子运动模式举例 (a) 原始轨迹中X 坐标随时间变化曲线; (b) 从第2 阶到第64 阶的连续小波变换系数Fig. A5. Example of lipid trajectories handled by wavelet:(a) X-coordinate profile of the original trajectory;(b) wavelet coefficients from the 2 nd to the 64 th.

图A6 第2 阶 (a)、32 阶 (b) 和64 阶 (c) 对应的小波变换系数; (d) 此轨迹经小波变换得到的第32 阶系数以及相应的阈值红线Fig. A6. The wavelet coefficients of the 2 nd, 32 th and 64 th scale; (d) the 32 th wavelet coefficient and the corresponding red line of threshold value.
图A7 (a) 第32阶系数小波变换区分后的轨迹曲线, 红色和蓝色分别对应轨迹中运动较快和较慢的阶段;(b) 与图 (a) 中红色、蓝色阶段相对应的MSD 分布Fig. A7. (a) Trajectory discriminated by wavelet using the 32 th scale coefficient, whose red part refers to the “fast” motion type and blue part refers to the “slow” motion type;(b)the corresponding MSD distributions of the red and blue parts.
选择置信区间为95% (a= 0.05),p为实验数据总体一致的概率,h为检验结果. 由表B2 中三组实验的结果分析, 扩散系数的分布大致相当,h为0, 实验结果可信.
3 脂分子扩散系数PDF 分布 (正文图2) 的统计价值讨论
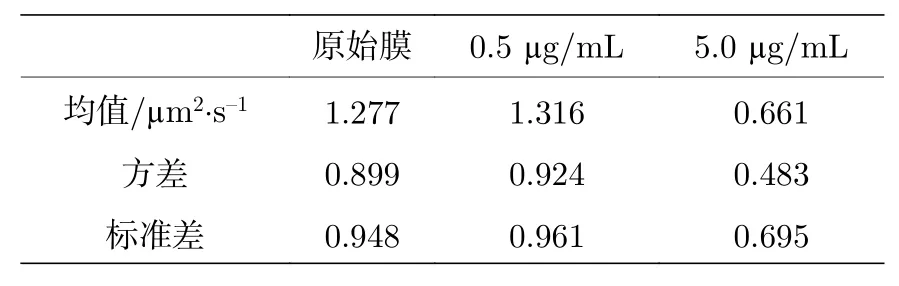
针对这个问题我们计算了每一个体系所有扩散系数的均值和方差 (标准差), 如表B3 所更. 由统计结果可以看出,方差 (标准差) 相对于均值显然是不可忽略的. 使用均值这一指标虽然可以反应体系的一些变化, 但是掩盖了体系中的一些关键的信息. 而如此之大的方差也意味着那些占比较小的部分不可忽略、具备统计价值.

表B1 实验样本数Table B1. Number of samples in different experimental conditions.

图A8 模拟体系中蜂毒肽加入前后脂质分子对应的MSD 分布. 蜂毒肽加入后, 脂质运动明显变慢Fig. A8. Changes in MSD distributions of lipids before and after the addition of Mel in the simulations.

表B2 基于三次独立实验结果的秩和检验置信度分析Table B2. Confidence analysis using rank sum test based on three independently repeated experiments.
表B3 脂分子扩散系数PDF 分布的方差分析Table B3. Variance analysis of the DL distribution in different conditions.
猜你喜欢
中成药(2018年9期)2018-10-09 07:18:36
中成药(2018年1期)2018-02-02 07:19:53
现代园艺(2017年13期)2018-01-19 02:28:09
中成药(2017年4期)2017-05-17 06:09:26
现代检验医学杂志(2016年3期)2016-11-15 01:59:28
药学与临床研究(2015年4期)2015-06-05 11:35:54
中国医疗美容(2015年4期)2015-04-27 02:24:11
科学中国人(2015年16期)2015-02-28 09:14:02
科学中国人(2015年13期)2015-02-28 09:13:26
食品工业科技(2014年23期)2014-03-11 18:18:53
