
岷山北坡高海拔区草本植物群落特征及主要植物功能群C、N、P含量分析
2020-05-29 09:06何靖田青宋玲玲
生态环境学报 2020年3期
何靖,田青,宋玲玲
甘肃农业大学林学院,甘肃 兰州 730070
随着人们对物种多样性及生态系统功能间关系的深入研究,植物多样性随环境因子变化的群落动态梯度、多样性以及生产力关系逐渐成为研究的热点(Fahrig et al.,2011;许传阳等,2014)。物种多样性沿海拔的变化规律是生物多样性研究的重要议题,海拔梯度涵盖了温度、湿度及太阳辐射等环境因素的综合影响,对物种多样性的垂直分布格局具有重要作用(Gaston,2000;Oommen et al.,2005)。同时,海拔差异影响水热条件的空间分布,进而影响区域植物群落的分布和结构(岳明等,2002)。环境因子影响植物的生物量,而海拔可以引起环境因子的垂直分异变化(李婷等,2015),从而引起环境因子的变化,继而影响植物生物量的大小。生物量是草地生态系统结构与功能的重要表现形式,是维持草地生态系统稳定发展的基础,对草地维护与管理的可持续发展起到重要作用(Tilman et al.,1996),其在叶、枝和根系间的分配是植物长期适应环境的结果,并在植物的生长、发育和繁殖等方面起重要作用(郝虎东等,2009)。作为森林生态系统特殊层片的林下草本群落,在保持水土、提高物种多样性以及促进物质循环和能量流动层面具有重要意义(张守昊等,2018)。此外,研究生态系统元素的平衡是当前全球变化生态学的热点和焦点(王绍强等,2008)。因此,在岷山北坡研究植物群落的海拔梯度格局以及物种多样性与植物营养元素的变化规律,对于理解草本群落生物量对海拔、物种多样性以及养分变化的响应具有重要意义。
试验区为岷山山系森林植被区,该区东接秦岭山地、西邻青藏高原、南近横断山脉和华中地区,其所在的甘肃白水江国家级自然保护区种子植物区系,汇集了中国——日本森林植物区、中国喜马拉雅森林植物区、青藏高原植物区三大植物区系成分,成分类型复杂多样,相对高差较大,具有浓缩的环境梯度和高度异质化的生境,气候和植被垂直分布明显,是岷山至秦岭物种基因交流和过渡的重要通道(田青等,2016)。目前,岷山北坡草本群落的研究主要侧重于生物量在海拔梯度上的分布特征(田青等,2016;张涛等,2019;罗立娇等,2019)和物种多样性对地形因子的响应(宋玲玲等,2017a;宋玲玲等,2017b)以及土壤养分的关系(张守昊等,2018),而对物种多样性随海拔的变化规律及功能群碳、氮、磷的相关研究较为缺乏。此外,对岷山北坡的研究区域主要集中在低海拔区,而对高海拔区的草本群落特征和物种多样性以及植物营养元素的研究较为缺乏。
本试验的研究对象为岷山北坡海拔 2300—2900 m的草本植物,对其进行实地调查和统计,同时测定地上生物量和地下生物量以及主要植物功能群的碳、氮、磷元素,分析了草本生物量的形成规律、物种多样性的变化特征,以及主要植物功能群碳、氮、磷元素在海拔梯度上的分布差异,为今后岷山北坡草本群落分布格局和生物地球化学循环的研究提供科学依据。
1 材料和方法
1.1 试验区概况
试验区选为岷山北坡两河口区域,该区域隶属于甘肃省白水江国家级自然保护区,地理坐标为104°19′—104°32′E,32°55′—32°94′N,海拔高度平均为1475 m,气温全年平均为15.6 ℃,降水量年均为840—950.3 mm,降水量主要集中在6—8月,为70%。该区植物群落的结构组成非常复杂,拥有从亚热带到暖温带再到中温带以及寒温带的多种代表性群落类型,其中海拔2300—2900 m区域的植被类型主要为针阔叶混交林带。基带土壤为山地暗棕壤,土体呈暗棕色或棕色,粘化作用强,呈弱酸性,pH值6.0—6.3,土壤有机质含量高且沿剖面变化和缓,土壤矿物质有弱脱硅富铝化作用存在。试验调查统计出木本植物15科37种,主要为红桦(Betula albosinensis)、糙皮桦(Betula utilis)、麻栎(Quercus acutissima)、麦吊云杉(Picea brachytyla)、岷江冷杉(Abies fargesii)、华椴(Tilia chinensis)、秦岭白蜡(Fraxinus paxiana)、元宝槭(Acer truncatum)、青榨槭(Acer davidii)、楤木(Aralia chinensis)、藏刺榛(Corylus ferox)等高大乔木,以及秦岭木姜子(Litsea tsinlingensis)、稠李(Padus racemosa)、山樱桃(Cerasus serrulata)、水栒子(Cotoneaster multiflorus)、锐齿臭樱(Maddenia incisoserrata)、陕甘花楸(Sorbus koehneana)、华北珍珠梅(Sorbaria kirilowii)、刺五加(Eleutherococcus senticosus)、蜡莲绣球(Hydrangea strigosa)、缺苞箭竹(Fargesia denudata)、桦叶荚蒾(Viburnum betulifolium)、金银忍冬(Lonicera maackii)等矮小乔木和灌木群落。草本植物共20科44种。主要草本植物见表1。
1.2 试验设计
试验于2014年8月中下旬在岷山北坡两河口海拔2300—2900 m地段进行,此时草本植物生长正处于旺盛期。试验采用样线和样地相结合的调查方法,且按照样线内海拔每升高大约100 m设置样地。样地的面积为20 m×20 m,在样地的四角和中间位置设1 m×1 m的草本样方5个。每块样地的海拔高度用GPS进行测定,并且记录样方内的所有植物种类数。
1.3 试验方法
统计小样方内所有的物种数和株数,同时记录植物的高度、盖度和密度。地上生物量的测定采用刈割法,齐地面刈割后在105 ℃下杀青10 min,然后在 70 ℃烘箱中烘至恒质量,称质量。地下生物量测定时先用土钻进行分层取样,然后清洗各土层钻取的土样,捡出所有根系,将其置于 80 ℃烘箱中烘至恒质量,称质量,最后换算为单位面积地下生物量(g·m-1)。参考已有植物功能群的分类标准(郭屹立等,2012),将测定的植物划分为禾本科、菊科和杂类草3个功能群,测定其有机碳含量(重铬酸钾硫酸溶液氧化法)、全氮含量(凯氏定氮法)和全磷含量(钼锑抗比色法)。
1.4 数据分析
用SPSS 19.0软件对草本植物的地上生物量、群落特征(包括盖度、物种丰富度、密度、高度、多度)的关系进行逐步回归分析,用 Duncan检验进行多重比较,最后用Pearson相关系数检验变量间的相关性。对草本植物功能群的 C、N、P与海拔梯度的关系采用曲线估计,并用SPSS软件的曲线估计得到P值。数据的处理用Excel 2010。
2 结果与分析
2.1 各海拔样地基本情况
共调查了7个样地,统计出草本植物20科35属 44种,其中,菊科、百合科和蔷薇科的物种数较多,分别占总物种数的25%,14%和11%。在海拔2338.2 m和2446 m处,植物类型主要为蔷薇科和莎草科,即东方草莓和苔草为优势种。随着海拔的升高,2539、2636.1及2731.9 m莎草科植物分布最多,即苔草成为主要的优势种。海拔继续升高,草本物种数和数量明显减少,而杂类草逐渐增多,且伞形科、鳞毛蕨科和唇形科植物取代蔷薇科和莎草科成为优势种(表1)。
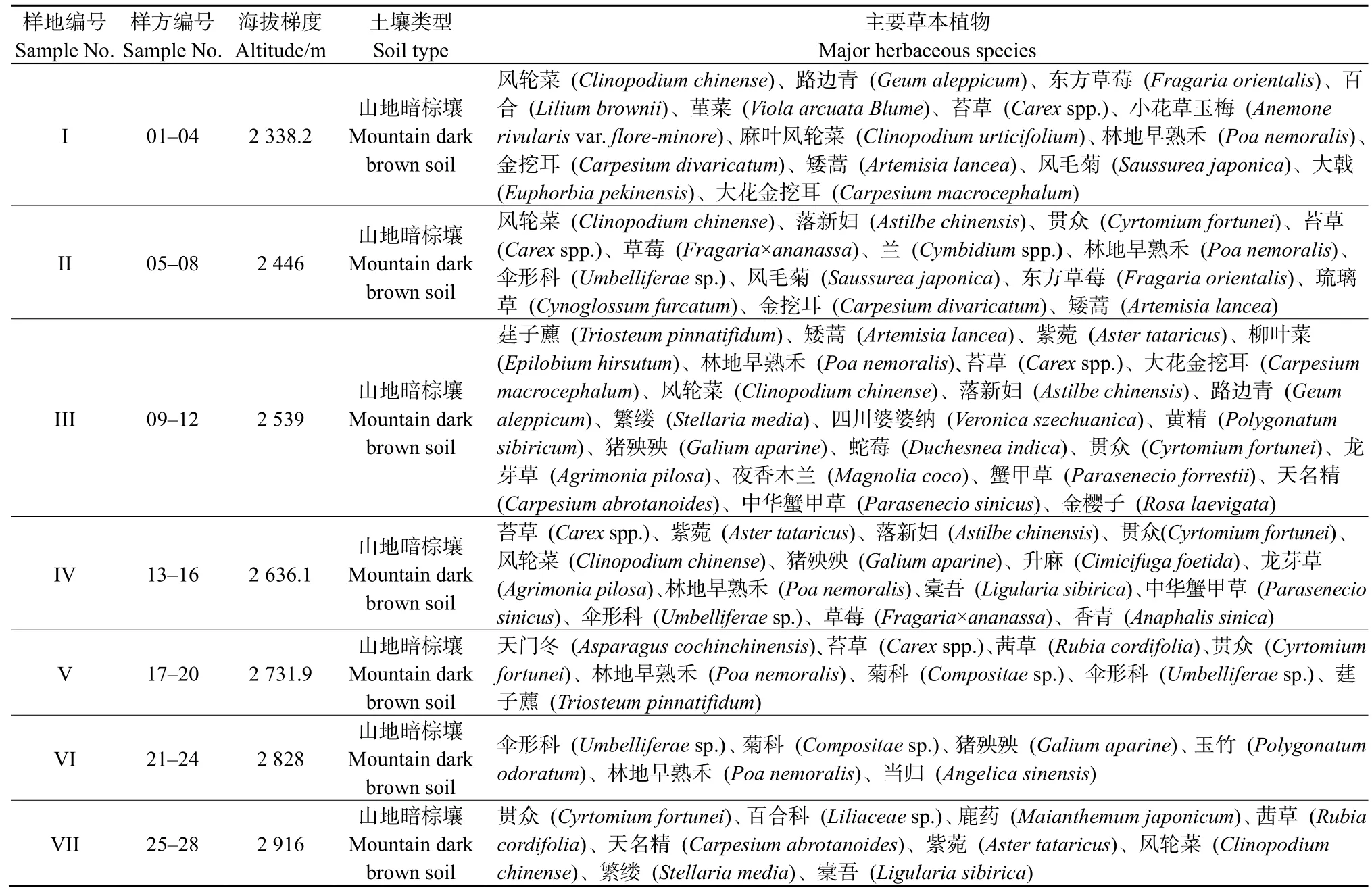
表1 草本植物的调查情况Table 1 Investigation of herbaceous plants
2.2 草本植物的生物量沿海拔梯度的变化规律
运用回归分析对草本植物生物量沿海拔梯度的变化规律进行比较,结果表现为地上生物量与海拔梯度的相关性不显著,其回归方程为:y=-0.0027x+29.806(R2=0.0003,P=0.925),地下生物量与地上生物量表现出一致的变化特征,即与海拔梯度的相关性也不显著,其回归方程为:y=-0.0037x+16.879(R2=0.0053,P=0.678)。海拔2636.1 m处,草本生物量与其他海拔相比差异较大,此时的值最大,而在海拔2828 m处时,生物量最小。地上、地下生物量随着海拔的不断升高都表现出先增加后减少的总体变化趋势(图1)。
2.3 不同海拔梯度草本植物群落特征

图1 不同海拔梯度草本植物群落生物量Fig.1 Biomass of herbaceous community in different altitude gradients
对草本植物的群落特征指标,包括盖度、物种丰富度、密度、高度、多度进行统计分析,结果显示,盖度在海拔2731.9 m处最大,为24.6%,最小值在海拔2828 m处,仅为2.5%;物种丰富度在海拔2539 m时最大为22,在海拔2828 m时最小为6;密度在海拔2446 m最大,其值为139.6 plant·m-2,海拔 2828 m和 2916 m时最小,其值都为 6.4 plant·m-2;植被高度在海拔2828 m处最大,为33.1 cm,在海拔2338.2 m最小为8.73 cm;多度在海拔2446 m处最大为53,在海拔2916 m处最小为3.56。
回归分析结果显示:盖度与海拔呈极显著负相关关系,回归方程为:y= -0.615x+14.082(R2=0.025,P<0.001);物种丰富度与海拔呈极显著负相关关系,回归方程为:y= -1.536x+18.429(R2=0.39,P<0.001);密度与海拔呈极显著负相关关系,回归方程为:y= -16.764x+112.91(R2=0.596,P<0.001);高度与海拔呈极显著正相关关系,回归方程为:y=3.52x+7.737(R2=0.655,P<0.01);多度与海拔呈负相关但不显著,回归方程为:y= -81.036x+549.71(R2=0.568,P>0.05)(图 2)。
2.4 植物群落特征相关性分析
对草本植物的生物量与群落特征进行相关分析(表2),结果显示:地上生物量与地下生物量、盖度均表现出极显著的正相关关系(P<0.01);地下生物量与盖度呈显著的正相关关系(P<0.05);密度与高度表现出显著负相关(P<0.05),与多度呈极显著正相关(P<0.01);高度与多度表现出显著的负相关关系(P<0.05);物种丰富度与其他各群落指标的相关性均不显著(P>0.05)。
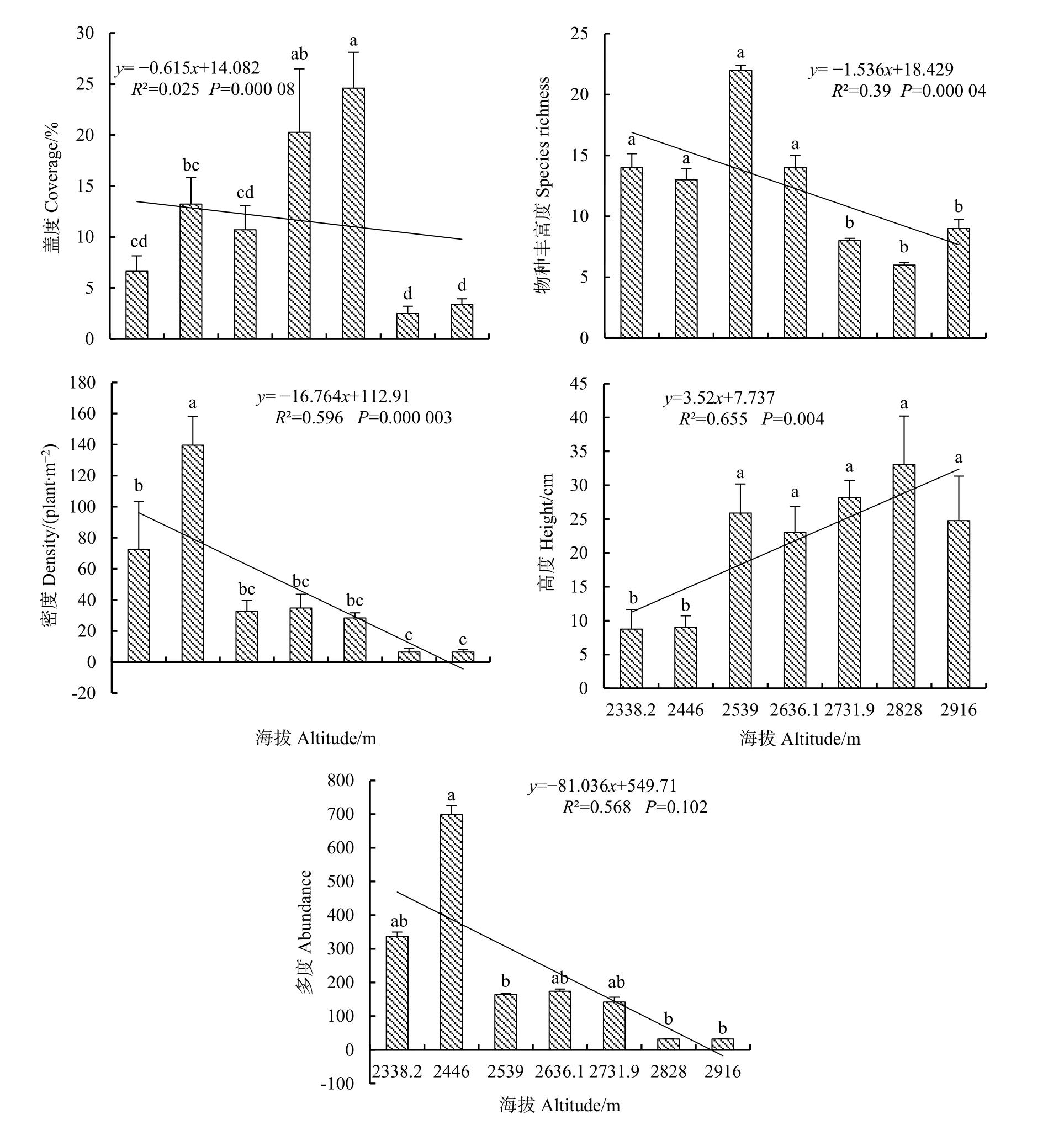
图2 不同海拔梯度草本植物群落特征Fig.2 Community characteristics of herbaceous plants in different altitude gradients

表2 植物群落特征相关性分析Table 2 Correlation analysis of plant community characteristics
2.5 草本植物功能群C、N、P分析
禾本科植物的叶碳含量(LCC)在海拔2828 m处出现最大值,叶氮含量(LNC)和叶磷含量(LPC)均在海拔2338.2 m处最大,而在海拔2446 m处,LCC、LNC、LPC都出现最小值。菊科植物的LCC在海拔2916 m处最大,LNC在海拔2731.9 m处最大,二者均在海拔2338.2 m处最小,而LPC在海拔2338.2 m时最大,在海拔2446 m处最小。杂类草植物的LCC在海拔2916 m处最大,在海拔2828 m处最小;LNC在海拔2731.9 m处出现最大值,在海拔2338.2 m处最小;LPC在海拔2539 m时最大,在海拔2446 m时最小。
从草本植物功能群的叶碳、氮、磷含量随海拔梯度的变化趋势来看,随着海拔梯度的不断升高,禾本科、菊科以及杂类草 3个植物功能群的 LCC整体呈上升趋势,LPC整体呈下降趋势,而 LNC禾本科呈下降趋势,菊科呈上升趋势,杂类草呈先上升后下降的变化趋势。相关分析表明:菊科植物的 LCC与海拔梯度呈极显著的正相关关系(R2=0.92841,P<0.01),其他LCC、LNC、LPC都与海拔梯度相关性不显著(P>0.05)(图3)。
禾本科的RCC在海拔2828 m处最大,在海拔2338.2 m处最小,而RNC在海拔2636.1 m处最小,RPC在海拔2446 m处最小,而RNC和RPC均在海拔2731.9 m处最大。菊科植物的RCC在海拔2916 m处最大,RNC在海拔2731.9 m处最大,二者均在海拔 2338.2 m处最小,而 RPC在海拔2539 m时最大,在海拔2446 m处最小。杂类草的RCC在海拔2916 m处最大,在海拔2338.2 m处最小;RNC在海拔2828 m处出现最大值,在海拔2636.1 m处最小;RPC在海拔2539 m时最大,在海拔2446 m时最小。
从草本植物功能群的根碳、氮、磷含量随海拔梯度的变化趋势来看,随着海拔梯度的不断升高,禾本科、菊科以及杂类草 3个植物功能群的 RCC整体呈上升趋势;禾本科和杂类草的 RNC呈上升趋势,而菊科表现出先上升后下降的趋势;禾本科的RPC呈上升趋势,而菊科和杂类草表现出先升后降的趋势。相关分析显示:菊科植物的 RNC与海拔呈显著的正相关关系(R2=0.83321,P<0.05),其他 RCC、RNC、RPC都与海拔梯度相关性不显著(P>0.05)(图4)(注:海拔2916 m没有调查到禾本科植物,故没有植物的C、N、P含量值)。
3 讨论
3.1 草本植物群落生物量在海拔梯度上的差异
生物量作为生态系统最基本的数量特征,反映了生态系统获取能量的能力,是研究草地生态系统物质循环,能量流动和生产力的基础(罗丹,2007)。同时,生物量也是测定群落结构和功能的主要指标,通常以地上生物量体现群落的结构特征和生长状况(Wang et al.,2005)。吴红宝等(2019)对藏北高寒草地物种多样性和生物量沿海拔的变化研究表明,地上生物量随海拔的升高表现出降低的变化趋势。本研究中,随着海拔的不断增加,草本生物量的变化趋势为先增加后减少,具体表现为地上生物量和地下生物量均在海拔 2636.1 m处最大,分别为64.129 g·m-2和 21.59 g·m-2,在海拔 2828 m 处都出现最小值,其值分别为 1.529 g·m-2和 0.475 g·m-2,且与海拔梯度相关性均不显著,这可能是由于研究区域的差异而导致的。有研究显示,群落的物种组成是其对环境长期适应的结果,并与环境形成了不同的小生物气候,影响了不同样地间草本生物量的差异性(岑宇等,2017),从而使得地上、地下生物量与海拔的相关性不显著。有研究表明,林分林龄和林分密度对林下植被生物量的变化影响较大(吴红宝等,2019)。林分从幼龄到成熟林过程中,其林冠层是一个未郁闭—郁闭—郁闭程度降低的动态过程,生物量则呈逐渐累积增加—逐渐减少—快速增加的变化过程(闫文德等,2003)。所以,上层乔木的树冠郁闭度及树形影响林下植被生物量的变化,这些因素通过影响落到林下草本层的光照来影响草本层的生长和发育,进而影响了林下草本群落的生物量(田青等,2016)。海拔在2636.1 m处时,林分的密度较小,导致林下植被的受光条件较好,使得草本植物生长较好,所以该区域生物量较大,而海拔2828 m处情况则相反,所以生物量较低。
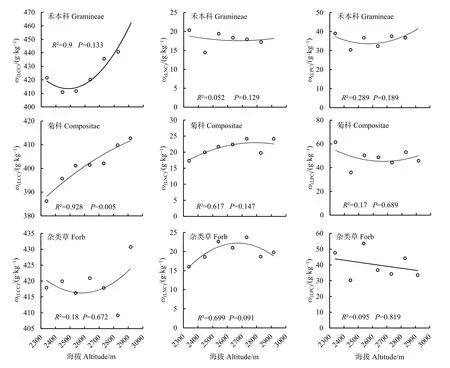
图3 不同海拔梯度草本植物功能群的叶碳、氮、磷含量特征Fig.3 Characteristics of leaf carbon,nitrogen and phosphorus contents in functional groups of herbaceous plants at different altitude gradients
3.2 海拔高度对草本群落物种多样性的影响
植物物种多样性及生物量的空间分布格局是各种生态因子作用的综合反映,海拔作为众多因子中最为关键的因素之一,其变化通常会影响温度、湿度及太阳辐射等的再分配,进而影响山地植物的分布格局(Gaston,2000;Li et al.,2011)。本研究结果显示,植物盖度与海拔梯度表现出极显著的负相关关系,这与吴红宝等(2019)对藏北高寒草地物种多样性以及王娟等(2019)对青藏高原草地植物群落的研究结果相一致。张丽霞等(2000)的研究表明,物种丰富度指数和物种多样性指数随海拔的升高逐渐减小,本研究显示,物种丰富度与海拔也呈极显著的负相关关系。密度与海拔表现出极显著的负相关关系,该结果与冯建孟等(2009)对地带性植物群落的物种多样性研究结果相一致。高度与海拔梯度呈极显著正相关关系,而田青等(2016)的研究结果显示,植被高度与海拔梯度无显著关系。本研究中,草本群落的生物多样性整体表现出中海拔较高,低海拔和高海拔较低的变化特征。有研究表明,低海拔区土壤温度高湿度低,抑制了植物的生长,高海拔区土壤湿度高温度低,而高湿度的环境不利于禾本科植物的生长(吴红宝等,2019),同时高海拔段风速急、土壤蒸发强及昼夜温差大(冯建孟等,2009)等恶劣条件只能使得适应力极强的物种生存,温度和水分的变化会引起物种组成的改变(Wu et al.,2011;Ganjurjav et al.,2016;Hopping et al.,2018)。因此,形成了中等海拔梯度物种多样性较高的分布格局。
3.3 不同海拔梯度草本植物功能群的C、N、P含量特征

图4 草本植物功能群的根碳、氮、磷含量与海拔梯度的关系Fig.4 Relationships between root carbon,nitrogen and phosphorus contents of herbaceous plant functional groups and altitude gradient
生态系统功能与物种数目关联的同时,也与物种所具有的功能特征相关(江小雷等,2016)。植物的根系固定着植物躯体、调节着植物的生长和发育、储存着职务所需的营养物质,可见对于植物形成地上生物量以及生长发育的整个过程起着重要作用(黄德青等,2011;胡中民等,2005)。N和P不仅对植物的个体发育和生理功能上起重要作用,还直接影响植物群落组成、结构和生态系统功能和生产力(Robertson et al.,1981;Vitousek et al.,1989)。本研究将草本植物按照功能群划分后,对其叶C、N、P含量和根C、N、P含量沿海拔梯度的差异进行分析,发现 3个功能群的叶 C、N、P含量和根 C、N、P含量的最值大致出现在相同的海拔高度处,比如禾本科植物的C、P含量,菊科植物的C、N含量,杂类草的C、P含量。有研究表明,磷含量的变化与海拔没有必然的联系,主要受到成土母质的影响(Shrestha et al.,2018),而本研究显示,随着海拔梯度的不断升高,禾本科、菊科以及杂类草3个植物功能群的LCC整体呈上升趋势,RCC也呈上升趋势,LPC呈下降趋势。杨思琪等(2017)的研究结果表明,随着海拔升高,LCC降低,LPC升高,说明在不同的研究区域,LCC和LPC存在差异。元素化学计量间的关系在一定程度上可以反映植物的生态策略,如C∶N和C∶P可以反映出植物的生长速度,同时N∶P可以指示植物的营养状况(罗立娇等,2019),在本研究中,菊科植物的LCC与RNC均与海拔梯度表现出显著的正相关关系。
4 结论
本研究利用回归分析揭示了草本群落生物量随海拔梯度的变化特征以及不同海拔梯度草本群落特征的差异,运用相关性分析探讨了草本植物地上、地下生物量与群落特征的相关性,最后用曲线估计分析了草本功能群的 C、N、P含量与海拔的关系。研究结果显示,草本群落的生物量均与海拔梯度的相关性不显著,且随海拔升高均表现出先增后减的总体趋势;植物盖度、物种丰富度、密度均与海拔表现出极显著负相关关系,而植被高度与海拔极显著正相关。随着海拔梯度的不断升高,禾本科、菊科以及杂类草3个植物功能群的叶碳含量整体呈上升趋势,叶磷含量整体呈下降趋势,而叶氮含量禾本科呈下降趋势,菊科呈上升趋势,杂类草呈先上升后下降的变化趋势。
猜你喜欢
散文诗(2021年22期)2022-01-12
青年文学家(2021年16期)2021-08-05
少儿美术(2020年8期)2020-11-06
环球时报(2019-05-23)2019-05-23
天然产物研究与开发(2018年10期)2018-11-06
现代园艺(2017年13期)2018-01-19
江苏农业科学(2016年3期)2016-05-03
山东农业科学(2014年8期)2014-10-10
小说月刊(2014年1期)2014-04-23
郑州大学学报(工学版)(2014年6期)2014-03-01
