
Standardized extract of Centella asiatica ECa 233 inhibits lipopolysaccharide-induced cytokine release in skin keratinocytes by suppressing ERK1/2 pathways
2020-05-29 07:06FuroidaMoolsapSupitaTanasawetMayureeTantisiraPilaiwanwadeeHutamekalinVaromyalinTipmaneeWanidaSukketsiri
Furoida Moolsap, Supita Tanasawet, Mayuree H.Tantisira, Pilaiwanwadee Hutamekalin, Varomyalin Tipmanee,Wanida Sukketsiri✉
1Department of Pharmacology, Faculty of Science, Prince of Songkla University, Songkhla, 90110, Thailand
2Department of Anatomy, Faculty of Science, Prince of Songkla University, Songkhla, 90110, Thailand
3Faculty of Pharmaceutical Science, Burapha University, Chon Buri Campus, Chon Buri, 20131, Thailand
4Department of Physiology, Faculty of Science, Prince of Songkla University, Songkhla, 90110, Thailand
5Department of Biomedical Sciences, Faculty of Medicine, Prince of Songkla University, Songkhla, 90110, Thailand
ABSTRACT
KEYWORDS: Centella asiatica; Cyclooxygenase-2; HaCaT;Interleukin-1β; Tumor necrosis factor-α
1.Introduction
The human largest organ in the body is the skin which possesses an important role in the immune defense mechanism.Nonetheless,an improper immune response results in the initiation of inflammatory skin disease[1].One of the most typical complications in dermatology is an inflammatory skin condition that affects the quality of life and produces excessive financial inconvenience.The inflammatory skin condition is characterized in acute condition by occasional rashes with itching as well as redness and in chronic condition elicited by dermatitis, rosacea, seborrheic dermatitis,and psoriasis[2].Upon skin injury, keratinocyte cytokines including tumor necrosis factor-alpha (TNF-α) and interleukin-1 beta (IL-1β)are produced responsible for cutaneous inflammation[3,4].To date,non-steroidal and steroidal anti-inflammatory drugs are common medications used to treat acute inflammation; however, they have not yet improved chronic inflammatory diseases.Additionally,gastrointestinal bleeding and cardiovascular complications are generally adverse effects found after treating with some of these medications[2].Recently, many studies have been conducted to suppress the inflammatory cytokines and chemokines of lipopolysaccharide (LPS)-inflamed keratinocytes and discover the linked mechanisms[5,6].Keratinocytes are the prominent cell type of epidermis which play an important part in the pathogenesis of inflammatory skin diseases such as atopic dermatitis[7].In order to represent the keratinocyte-like features, HaCaT cell line which is an immortalized aneuploidy from normal adult human skin has been employed in the in vitro study[8].
In traditional medicine, Centella asiatica (C.asiatica) (Linn)Urban has been generally consumed for treating a wide range of dermatological diseases[9] that contains a source of various bioactive compounds including asiaticoside, asiatic acid, madecassoside and madecassic acid[10,11].C.asiatica possesses excellent antioxidant and antiinflammatory properties[12,13].C.asiatica has been reported to suppress inflammatory responses in macrophages[12,13] and attenuate phthalic anhydride-induced atopic dermatitis in mice[13].Madecassoside and asiaticoside, a major constituent isolated from C.asiatica, have also been extensively investigated on its pharmacological activities.Asiaticoside and madecassoside have potent anti-inflammatory effects in vitro[14] and protective effects against LPS-induced inflammation in an animal model[15-18].ECa 233, a C.asiatica extract with the patent-pending procedure, was kept the ratio between madecassoside and asiaticoside in a range of 1.5 ± 0.5 and also ensured the content of triterpenoid glycoside at least 80%[19].Since ECa 233 displays anti-inflammatory activity[20],it is likely to note that this compound could promote skin disorder treatment.Nonetheless, the cellular mechanism of ECa 233 in ameliorating LPS-induced inflammatory cytokines in keratinocytes has not been clarified.The aim of this study was to evaluate the antiin flammatory properties of ECa 233 on the inflammatory conditions in human keratinocytes.
2.Materials and methods
2.1.Cell culture and reagent
Human keratinocytes (HaCaT) cell line was obtained from the CLS cell line service (Heidelberg, Germany).HaCaT cells were maintained in Dulbecco’s modified Eagle’s medium (Biochrom,Germany) containing 10% fetal bovine serum, 1% antibiotic(penicillin/streptomycin) and 1% L-glutamine (Gibco, USA) in a CO2incubator at 37 ℃.ECa 233, a standardized C.asiatica extract,was provided by Siam Herbal Innovation Co., Ltd (Samutprakan,Thailand).Liquid chromatography-tandem mass spectrometry was used to quantify the contents of madecassoside (52%) and asiaticoside (41%) in the standardized C.asiatica extract ECa 233[19,21].ECa 233 was dissolved in dimethylsulfoxide (Sigma,St.Louis, MO, USA).Dimethylsulfoxide concentration in culture medium was consistently less than 0.1% (v/v), which was non-toxic to the HaCaT cells.Lipopolysaccharide (LPS) from Escherichia coli 055:B5 was obtained from Sigma (St.Louis, MO, USA).Other reagents were all of high grade for experiments.
2.2.Determination of nitric oxide (NO) level and cytotoxicity in HaCaT cells
In our previous study, ECa 233 (0.1-100 μg/mL) did not cause a reduction in viability of HaCaT cells after 24 h treatment[22], so we used this concentration for all experiments in this study.HaCaT cells were activated with 10 μg/mL of LPS and then administered with 0.1, 1, 10 and 100 μg/mL of ECa 233 for 24 h.Additionally,we used either 100 μg/mL of ECa 233 or 1 μg/mL dexamethasone alone in order to observe the individual effect of ECa 233 and dexamethasone on HaCaT cells.After incubation, Griess reagent was used to evaluate nitrite levels in the medium.In brief, 100 μL of Griess reagent was combined with 100 μL of media and incubated for 10 min.Then, a microplate reader (Bio-Tex Instruments, Inc.,VT, USA) was utilized to measure the nitrite-containing samples in culture medium at 540 nm.A standard curve was made using sodium nitrite.In addition, the cells were incubated with 500 μg/mL of MTT solution for 2 h and then the HaCaT viability was detected at 570 nm by using a microplate reader.
2.3.Evaluation of intracellular reactive oxygen species (ROS)generation in HaCaT cells
The oxidation of 2′,7′-dichlorofluorescein diacetate (Sigma, St.Louis, MO, USA) was used to assay the production of intracellular ROS in HaCaT cells.Briefly, HaCaT cells were grown in 96-well plate at the density of 5×104cells/well.Then, HaCaT cells were stimulated with LPS in the presence or absence of the ECa 233 at 0.1, 1, 10 and 100 μg/mL for 24 h.Dexamethasone (Sigma,St.Louis, MO, USA) at the concentration 1 μg/mL was used as a reference drug.After 24 h of incubation, the cells were exposed with 50 μM of 2′,7′-dichloro fluorescein diacetate for 1 h, at 37 ℃ in dark condition.A fluorescence microplate reader (Bio-Tex Instruments,Inc., VT, USA) was used to assess the intensity of DCF at 485 nm of excitation wavelength and 530 nm of emission wavelength.
2.4.Determination of prostaglandin E2 (PGE2), TNF-α and IL-1β production
IL-1β and TNF-α have been stated to play a critical role during inflammatory responses[23,24].HaCaT cells were stimulated with LPS in the presence or absence of 1 μg/mL dexamethasone and ECa 233 at concentrations of 0.1, 1, 10 and 100 μg/mL for 24 h.After 24 h incubation, the ELISA method was used to evaluate the level of PGE2, TNF-α, and IL-1β in the cultured medium of HaCaT cells.
2.5.Determination of the expression of cyclooxygenase-2(COX-2), extracellular signal-regulated kinase 1/2(ERK1/2) and nuclear factor-κB (NF-κB) in HaCaT cells
After the cells were stimulated with LPS in the presence or absence of ECa 233 (0.1, 1, 10 and 100 μg/mL) and 1 μg/mL of dexamethasone for 24 h, RIPA buffer contained protease and phosphatase inhibitor cocktail was applied to HaCaT cells at 4 ℃ for 30 min for cell lysis.Bradford protein assay (Bio-Rad, California,USA) was utilized to determine the concentration of protein in each collected sample.Then, proteins were loaded to 10% SDS-polyacrylamide gel electrophoresis and moved to nitrocellulose membranes.Then, the membranes were blocked using 5% skim milk in Tris-buffered saline, and 0.1% Tween 20.Then, primary antibodies against NF-κB (Santa Cruz Biotechnology; 1:200),ERK1/2 (Abcam; 1:1 000), p-ERK1/2 (Abcam; 1:1 000), and COX-2(Thermo Scientific; 1:1 000) were incubated overnight at 4 ℃.Then,the secondary antibody conjugated with HRP (Thermo Scientific,Waltham, Massachusetts, USA) was applied for 1 h at room temperature.Chemiluminescence (Supersignal West Pico; Pierce Biotechnology) was used to recognize the specific protein bands.
2.6.Immunofluorescence
After the cells were stimulated with LPS in the presence or absence of ECa 233 and dexamethasone for 24 h, the HaCaT cells were fixed with 4% paraformaldehyde for 20 min and soaked twice with PBS.Subsequently, the HaCaT cells were incubated with 0.1% Triton X-100 for 20 min and blocked nonspecific proteins with bovine serum albumin (1%) for 1 h.Then, the cells were incubated with the NF-κB primary antibody (Santa Cruz Biotechnology) (1:100)at 4 ℃ overnight, followed by incubation with FITC conjugated secondary antibodies for 1 h at room temperature.The nuclei were counterstained with Hoechst 33342 (10 μg/mL).The cells were then identified under a fluorescence microscope (Olympus IX73, Japan).
2.7.Statistical analysis
All data were represented as mean ± SD.One-way analysis of variance (ANOVA) was used for testing the differences of the mean values among groups and pairwise comparison between groups was analyzed by LSD post hoc test.P<0.05 was considered statistically significant.
3.Results
3.1.Effect of ECa 233 on LPS-induced ROS and NO levels in HaCaT cells
As shown in Table 1, the level of intracellular ROS significantly raised approximately 134.76% of untreated control when the keratinocytes were treated with LPS alone.After 10 and 100 μg/mL ECa 233 treatment, ROS level was significantly decreased to 121.83% and 116.91%, respectively (Table 1).In contrast to the cells stimulated with LPS, treatment with ECa 233 at concentrations of 0.1, 1, 10 and 100 μg/mL and 1 μg/mL dexamethasone for 24 h did not significantly change the content of nitrite, as a NO indicator(Table 1).Additionally, the inhibition was not due to chemically induced cytotoxicity at any dosages that were measured by MTT(Figure 1).
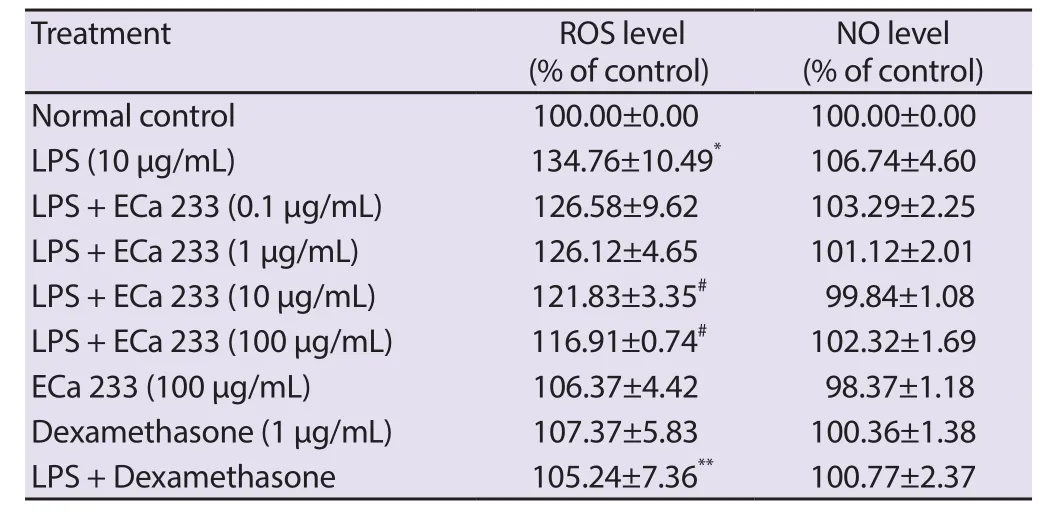
Table 1.Effect of standardized extract of Centella asiatica ECa 233 on ROS generation and NO level in LPS-inflamed skin keratinocytes.
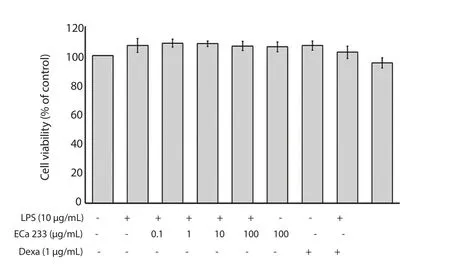
Figure 1.Effect of ECa 233 on the cell viability in LPS-inflamed skin keratinocytes.Data are represented as mean ± SD of four independent experiments.Dexa: dexamethasone.LPS: lipopolysaccharide.
3.2.Effect of ECa 233 on LPS-induced PGE2 level and COX-2 expression
As shown in Figure 2A-2C, the PGE2level and the expression of COX-2 protein were significantly increased in the LPS-stimulated HaCaT cells.Interestingly, ECa 233 (1, 10 and 100 μg/mL) and dexamethasone treatments showed a significant reduction in the level of PGE2in the HaCaT cells.In addition, dexamethasone and all concentrations of ECa 233 could inhibit the expression of COX-2 protein in the LPS-stimulated keratinocyte cells.The treatment of ECa 233 and dexamethasone alone did not affect the PGE2level and COX-2 protein expression.Based on this finding, it was observed that ECa 233 was responsible for the anti-inflammatory activity via the reduction of COX-2 protein expression and subsequently decrease of PGE2generation.
3.3.Effect of ECa 233 on LPS-activated TNF-α and IL-1β levels
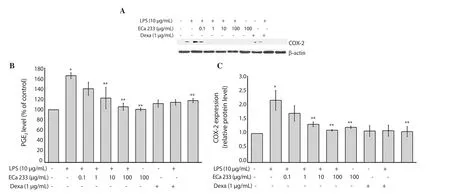
Figure 2.Effect of ECa 233 on PGE2 level and COX-2 expression in LPS-inflamed skin keratinocyte HaCaT cells.(A) COX-2 expression; (B) PGE2 level was evaluated by using ELISA; (C) Relative protein level of COX-2.Data are expressed as mean ± SD of four independent experiments.*P<0.001 compared to the untreated control, **P<0.001 compared to LPS alone.Dexa: dexamethasone.LPS: lipopolysaccharide.
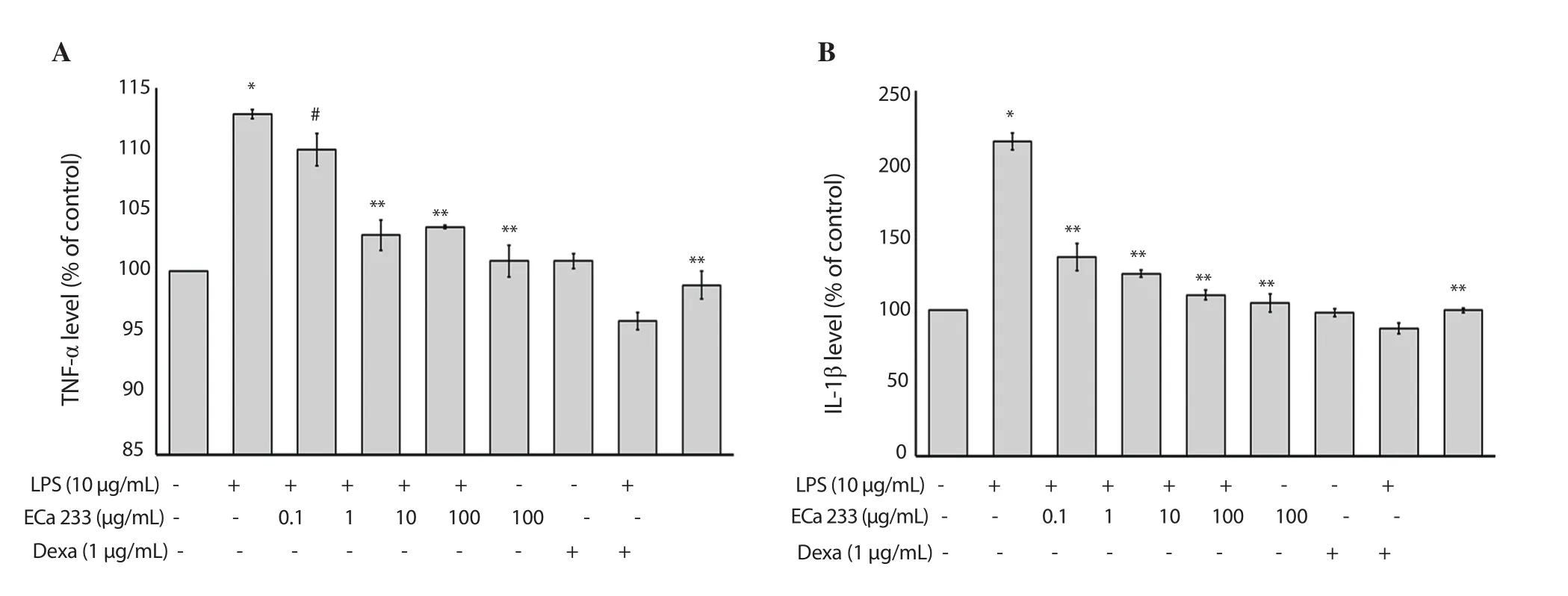
Figure 3.Effect of ECa 233 on TNF-α and IL-1β in LPS-inflamed skin keratinocyte HaCaT cells.TNF-α and IL-1β levels were determined using an ELISA method.(A) TNF-α level and (B) IL-1β level are represented as mean ± SD (n = 4).*P<0.001 compared to the untreated control, #P<0.05 compared to the LPS alone group, **P<0.001 compared to the LPS alone group.Dexa: dexamethasone.LPS: lipopolysaccharide.
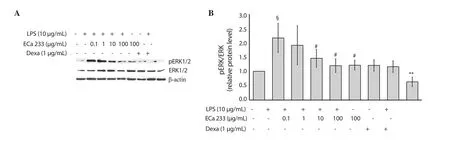
Figure 4.Effect of ECa 233 on LPS-induced inducible ERK1/2 expression in skin keratinocyte HaCaT cells.(A) the gel photographs display ERK1/2 protein expression, (B) the relative protein ratio of pERK/ERK is represented as mean ± SD (n=4).§P<0.01 compared to the untreated control, #P<0.05 compared to the LPS alone group, **P<0.001 compared to the LPS alone group.Dexa: dexamethasone.LPS: lipopolysaccharide.
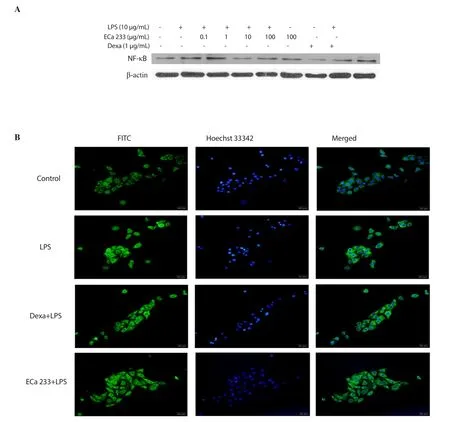
Figure 5.Effect of ECa 233 on p65 NF-κB expression and translocation in LPS-inflamed skin keratinocyte HaCaT cells.(A) NF-κB was examined by Western blotting assay and (B) by fluorescent microscopy after being stained with the NF-κB primary antibody, followed by incubation with FITC conjugated secondary antibodies.Dexa: dexamethasone.LPS: lipopolysaccharide.
As shown in Figure 3A and 3B, the levels of IL-1β and TNF-α were significantly enhanced after treatment with LPS at a concentration of 10 μg/mL in HaCaT keratinocytes.All concentrations of ECa 233 and dexamethasone ameliorated these changes in LPS-stimulated HaCaT cells (Figure 3A and 3B).Thus, these results indicated that LPS-induced skin inflammation could be alleviated by ECa 233 through terminating the production of proinflammatory cytokines.
3.4.Effect of ECa 233 on ERK1/2 expression
The ERK1/2 pathway involvement in the LPS-activated skin inflammation was investigated.The expression of phospho-ERK1/2 was significantly upregulated after 24 h of LPS treatment without changing the expression of total ERK1/2 protein in keratinocytes HaCaT (Figure 4A and 4B).Interestingly, the expression of phospho-ERK1/2 was significantly decreased after ECa 233 (1, 10 and 100 μg/mL) and dexamethasone treatments in the LPS-induced keratinocytes HaCaT (Figure 4A and 4B).Additionally, ECa 233 and dexamethasone alone did not induce alteration of the ERK1/2 phosphorylation.These results confirmed ECa 233 showed the anti-inflammatory activities against LPS-activated inflammation in keratinocytes partly via suppression of the ERK1/2 signaling pathway.
3.5.Effect of ECa 233 on the activation of NF-κB in keratinocyte HaCaT cells
To investigate the ECa 233 anti-inflammatory mechanism, we examined its effect on the activation of NF-κB in LPS-induced HaCaT cells.As shown in Figure 5A, the expression of NF-κB p65 protein was enhanced in LPS-stimulated keratinocytes.After ECa 233 and dexamethasone treatments, the level of NF-κB expression was decreased.Moreover, the nuclear translocation of NF-κB p65 was determined by immunofluorescence assays and shown in Figure 5B.In immunofluorescence assays, 10 μg/mL of ECa 233 was used because there was no significant difference on the potency between 10 and 100 μg/mL as shown in all results previously(Figure 1 to Figure 4).Therefore, low dose was used due to its low toxicity.The results showed that LPS-activated keratinocyte HaCaT cells presented larger numbers of nuclear NF-κB p65 than untreated HaCaT cells (Figure 5B).Dexamethasone and 10 μg/mL ECa 233 could slightly reduce the intensity of NF-κB p65 nuclear staining, illustrating that ECa 233 had abolished NF-κB p65 nuclear translocation in LPS-inflamed keratinocytes HaCaT cells (Figure 5B).These results indicated that ECa 233 might suppress the stimulation of NF-κB signaling pathway.
4.Discussion
Over the past decade, traditional herbal extracts are capable to abolish inflammatory disorders[25].C.asiatica is a source of natural bioactive constituents, triterpenoid saponins, flavonoids, phenolic acids and triterpenic steroids[10,11] and also used in the therapy of dermatological conditions including small wounds, hypertrophic wounds, burns, atopic dermatitis and psoriasis[9].ECa 233 has been reported as a standardized extract from C.asiatica containing a high level of asiaticoside and madecassoside[19,21].Remarkably, C.asiatica extract and its bioactive components have vigorous antiin flammatory effects in vitro[14-18,20] and protective effects against chemical-induced inflammation in animal model[15-18].Moreover,C.asiatica was used as an active compound in skin care due to its antioxidant, anti-inflammatory, and anti-aging activity[9,13].
Previous study has been reported on the association between LPS-induced inflammations and the activation of keratinocytes.Concurrent with this process, active keratinocytes produce the cytotoxic-inflammatory mediators containing reactive oxygen and nitrogen intermediates, hydrolytic enzymes, and inflammatory cytokines[26].Inflammatory state has been triggered by either NO or ROS[27].Oxidative stress is elevated in patients with skin inflammatory-associated diseases[28].We found that the generation of intracellular ROS was enhanced in LPS activated cells compared to the untreated cells, which was suppressed by 10 and 100 μg/mL of ECa 233 in human keratinocyte HaCaT cells.The inhibitory effect was not due to the cytotoxicity of ECa 233.This result is consistent with previous studies, implying that C.asiatica, ECa 233 and its active components possess antioxidant property in both in vitro and in vivo model[19,29-32].
Additionally, it has been known that the conversion of PGE2from arachidonic acid is catalyzed by COX-2.Several studies proposed that the elevated level of PGE2and the activity of COX may develop skin inflammation[33].In addition, the high level of COX-2 expression is related to the skin barriers disruption[34].In this study, our results showed that ECa 233 treatment at 1, 10 and 100 μg/mL caused a reduction of PGE2production and COX-2 protein expression in LPS-inflamed HaCaT cells.This result agrees with the previous finding which demonstrated C.asiatica extract,ECa 233 and its active components significantly decreased the PGE2production and COX-2 expression in both in vitro and in vivo study[13,17,20].Taken together, these data illustrate that the antiin flammatory mechanisms of ECa 233 are linked to the control of COX-2 protein expression in keratinocytes.
Keratinocyte was investigated as the primary source of epidermis cytokines and stated to release cytokines including IL-1, IL-3,IL-6, IL-8, colony-stimulating factor, TNF-α, transforming growth factor (TGF) alpha (TGF-α) and TGF-β[35].TNF-α and IL-1β are the most dominant secreted cytokines that negotiate and involve in the skin inflammatory disorders including psoriasis, and atopic dermatitis[24,25].TNF-α plays a crucial role in proinflammatory cytokines cascade and thereby results in the development of inflammation[36].IL-1β is involved in the progression of pathophysiology that develops during skin inflammation including dermatitis, and psoriasis[37].Therefore, the upregulation of proinflammatory cytokines’ mediators in the skin area indicates the inflammatory skin disorders[35].Our present study demonstrated that ECa 233 suppressed LPS-stimulated TNF-α and IL-1β production.Specifically, the secretion of IL-1β was more susceptible to be suppressed by ECa 233 than TNF-α.Several reports revealed that ECa 233 and its active compound derived from C.asiatica could decrease the proinflammatory cytokine including TNF-α, IL-1β, IL-6, and interferon gamma in both in vitro and in vivo model through suppressing NF-κB signaling pathways[13,14,16,20].The results give a hint that TNF-α and IL-1β would be a crucial factor for the antiinflammatory activity of ECa 233 in skin keratinocytes.
The transcription of inflammatory negotiators such as inducible nitric oxide synthase, COX-2, IL-1β, IL-6, and TNF-α is mediated by the control of NF-κB and the mitogen-activated protein kinases signaling pathway that is influenced during the progression of acute and chronic inflammatory diseases[38,39].Stimulation of the NF-κB signaling cascade by LPS and other cytokines, IκB proteins are phosphorylated by IκB kinase, then ubiquitinated and additionally impaired by the proteasome[40].Thereafter, an active form of NF-κB is secreted and changed to activate the target gene transcription in the nucleus[38].In keratinocytes, the activation of NF-κB is affected in the progression of skin inflammation and associated with the production of inflammatory cytokines.In addition, activation of ERK1/2 leads to increased production of proinflammatory mediators,such as TNF-α and IL-1β[39].Previous reports showed that NF-κB and ERK1/2 signaling pathways are affected by the control of cytokine, chemokine, and adhesion molecule generation in HaCaT keratinocyte cells[41,42].In this present finding, we demonstrated that ECa 233 inhibited the expression of NF-κB p65 determined by Western blotting and immunocytochemistry.Moreover, the upregulation of LPS-induced phosphorylation of ERK1/2 was also significantly suppressed by ECa 233.These results demonstrate that ECa 233 could inhibit the production of TNF-α, IL-1β, and PGE2and also the expression of COX-2 by suppressing the activation of ERK1/2 pathways.
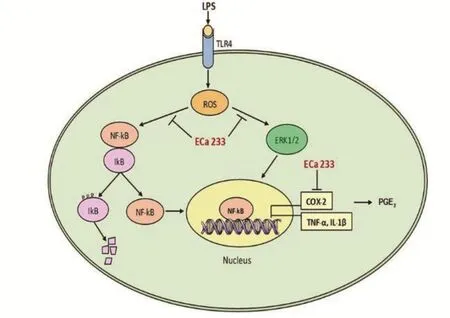
Figure 6.A possible mechanism of ECa 233 inhibiting LPS-inflamed skin keratinocyte.
ECa 233 exerts a significant anti-inflammatory effect via the decline of PGE2generation, which is due to the inhibition of COX-2 expression, as well as of proinflammatory mediators, including IL-1β and TNF-α.ECa 233 inhibited ERK1/2 activation through the suppression of the release of proinflammatory cytokines in LPS-stimulated skin inflammation (Figure 6).In conclusion, our findings propose a new therapeutic usage of ECa 233 towards skin inflammation.
Conflict of interest statement
We declare that there is no conflict of interest.
Funding
The accomplishment of this study was supported by grant from the General Project and Invention of Prince of Songkla University(SCI600421S) and Graduate School of Prince of Songkla University,Songkhla, Thailand.
Authors’ contributions
FM and WS designed the experiment, performed data analysis and statistics, and drafted the manuscript.ST, MHT, PH and VT interpreted the results and provided professional comments for this work.All authors have read and approved the final manuscript.
 Asian Pacific Journal of Tropical Biomedicine2020年6期
Asian Pacific Journal of Tropical Biomedicine2020年6期
- Asian Pacific Journal of Tropical Biomedicine的其它文章
- Enzyme-treated date plum leave extract ameliorates atopic dermatitis-like skin lesion in hairless mice
- Leishmania tropica: The comparison of two frequently-used methods of parasite load assay in vaccinated mice
- Formononetin alleviates diabetic cardiomyopathy by inhibiting oxidative stress and upregulating SIRT1 in rats
- Moringa oleifera leaf ethanol extract ameliorates lead-induced hepato-nephrotoxicity in rabbits
- Response surface methodology-based optimization of ultrasound-assisted extraction of β-sitosterol and lupeol from Astragalus atropilosus (roots) and validation by HPTLC method