
气温变化对温带混交林生态系统CO2通量的影响1)
2020-05-29 01:27龚元张银龙
东北林业大学学报 2020年5期
龚元 张银龙
(南方现代林业协同创新中心(南京林业大学),南京,210037)
在全球气候变化的背景下,地表城市化过程、人类工业活动、化石燃料使用以及世界人口的增加,导致大气中二氧化碳(CO2)、甲烷(CH4)等温室气体浓度上升,带来了诸多环境问题(主要是温室效应)。人们开始关注CO2、CH4等温室气体的排放动态的研究[1-5]。
森林和草地生态系统是陆地生态系统的主要碳汇[6]。植被的光合作用会吸收大气中部分CO2[7-10],由于植物自身独特的生长特性,植物生长状况及生理活动对气候变化的响应,影响森林、草地生态系统的碳循环过程。因此,开展森林、草地生态系统碳循环过程的研究,对分析全球碳循环过程,应对和解决全球气候问题具有重要作用[11-15]。
20世纪90年代以来,随着涡动相关(涡动协方差)技术(EC)的应用,为直接观测不同生态系统CO2排放和吸收动态提供了技术方法[5]。涡动相关技术作为观测一定生态尺度内下垫面与大气间物质循环和能量流动的技术手段,早期多应用于森林、湿地、草地等自然生态系统与大气间CO2交换的研究[16-22]。为了更好的研究陆地生态系统碳循环过程,涡动相关技术在森林生态系统CO2通量应用的研究成为热点。为了全球多站点通量数据的整合分析和通量数据处理标准化,形成了国际通量网(FLUXNET)以及中国通量网等区域-全球的通量观测数据共享网络[22-27]。国际通量网是较大的通量数据共享平台,目前注册通量观测站点已经超过900个,有相当数量的森林生态系统通量观测站分布在温带地区,包括较早开展通量观测的美国马萨诸塞州哈佛森林站点[21-22]。运用FLUXNET发布的通量数据,对全球森林生态系统CO2交换的研究也较多,主要包括:森林生态系统CO2通量动态特征、通量源区足迹的气候态特征、CO2通量建模等方面[17,21]。其中CO2通量建模是森林生态系统CO2通量研究的重要内容,主要包括CO2通量预测和CO2通量物候特征的提取等[17]。森林生态系统CO2通量物候特征(CO2通量变化量特征)为了解CO2通量对气候变化的响应提供了帮助。以往森林生态系统物候特征的提取多基于遥感数据中的植被产品(如NDVI、EVI和GPP等),计算模型多使用非对称高斯函数、D-L拟合和S-G滤波法等[28-32]。随着涡动相关系统的推广,高精度和高时间分辨率的CO2通量观测数据为森林生态系统CO2通量建模提供了高质量的数据支撑。Gu et al.[17]发展了基于涡动相关的植物群落光合作用参数化物候模型,该模型基于年际尺度CO2通量数据,拟合和提取CO2通量的物候特征。Richardson et al.[13]使用物候模型分析了国际通量网多站点生态系统净CO2交换(NEE)和总初级生产力(GPP)的物候特征及其与气候变化的关系,认为常绿针叶林的生产力对物候的敏感性较低;Yi et al.[16]将物候模型运用到草原通量站点,认为该模型具有分析长时间序列气候变化与碳反馈机制的研究;Niu et al.等[15]使用国际通量网CO2通量数据和物候模型,分析了北半球生态系统CO2通量对年均气温变化的响应,认为68个通量站点的物候特征均对年均气温变化的敏感性较高。
研究森林生态系统碳循环过程及其对气温变化的响应,可以为进一步了解陆地生态系统碳循环机制,为合理布局森林结构提供参考。该研究使用国际通量网所提供的FLUXNET2015数据集,即美国马萨诸塞州哈佛森林通量观测站点(LTER,US-Ha1)2000—2012年的CO2通量和气温观测数据等[21-22],基于植物群落光合作用的物候模型,分析温带森林生态系统CO2通量的变化量特征及其对气温变化的响应。
1 研究区概况
该研究区位于美国马萨诸塞州皮特舍姆市的哈佛林区,该区域所属温度带为温带,气候为温带大陆性湿润气候。定位环境观测塔(EMS)的地理坐标为(42.53°N,72.17°W),位于哈佛森林长期生态观测研究站内,通量观测塔搭载涡动相关通量观测系统和梯度微气象观测系统,塔高距离地面30 m(高过冠层高度5 m),海拔340 m,当地风向多为西南风和西北风。通量观测站点由美国哈佛大学所建立,从1988年开始对哈佛森林进行气象、生态环境等观测研究,该森林站点在FLUXNET的注册ID为US-Ha1[21],积累了较长时间尺度的通量数据和微气象数据,还包括地上生物量、凋落物、土壤温湿度、叶面积指数等生态数据[21]。该区域多年平均气温为6.62 ℃,其中最高月均温出现在7月份,最低月均温出现在1月份;多年平均年降水量为1 071 mm(见表1)。该区域下垫面多为木本植物,主要包括:北方红橡木(Quercusrubra)、红枫(Acerrubrum)、铁杉(Tsugacanadensis)等落叶阔叶林,平均冠层高度约20~24 m[21]。由于受到观测高度、风速和风向等因素的影响,该区域的通量源区范围可延伸至上风向1 km处[21]。
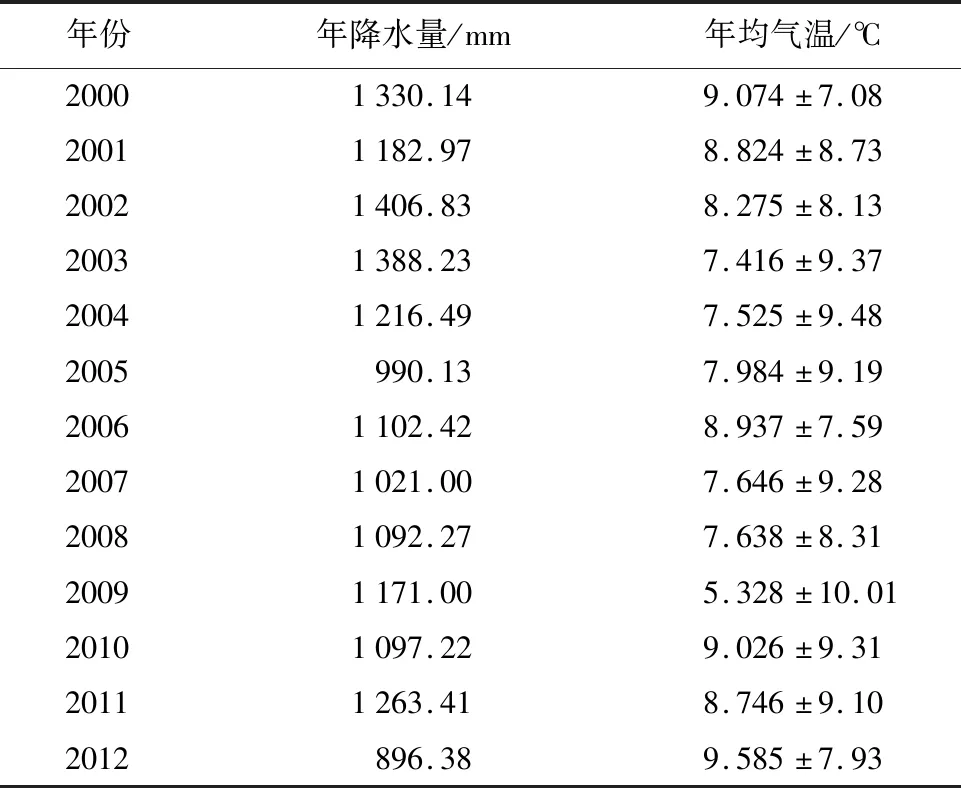
表1 2000—2012年哈佛森林生态系统年降水量和年均气温
2 研究方法
2.1 CO2通量与气温数据来源
该研究所使用的CO2通量和气温数据为国际通量网(FLUXNET)提供的美国哈佛森林长期生态观测站点(US-Ha1)观测和记录的2000—2012年的CO2通量数据和气温数据(Ta)。其中CO2通量数据已剔除野点、坐标轴旋转、密度修正等通量数据预处理;CO2通量数据主要包括生态系统净CO2交换(NEE)和总初级生产力(GPP)[21-22]。当NEE为正值时,代表生态系统处于释放CO2的状态,即碳源,当NEE为负值时,代表生态系统处于吸收CO2的状态,即碳汇;GPP为单位时间和单位面积内植物通过光合作用所固定有机碳的总量,也是衡量植被碳汇强度和研究植被碳循环的重要指标。
2.2 参数化植物群落光合作用物候模型
为了分析哈佛森林2000—2012年NEE和GPP的变化量特征,以及讨论CO2通量对气温变化的响应,参考生态系统总初级生产力(GPP)的植物群落光合作用物候模型[17],该模型运用年际尺度(1~365 d)每天每半小时最大GPP量化植物群落光合能力的季节变化。因此,本研究将物候模型应用于哈佛森林生态系统NEE、GPP和气温的变化量的计算,分析CO2通量和气温的变化量特征。将NEE、GPP变化量的最大点、0点和最小点出现的日序,与气温变化量的最大点、0点和最小点出现的日序进行线性拟合,分析CO2通量对气温变化的响应。
3 结果与分析
3.1 生态系统CO2通量观测值特征
由表2可知,2000—2012年的NEE年总量的最大值为298.13 g·m-2·a-1,出现在2010年,其次为60.86 g·m-2·a-1,出现在2011年,说明哈佛森林生态系统在2010年和2011年表现为碳源;在其它年份的年NEE总量均为负值,即表现为碳汇,其中在2008年的碳汇强度最大。GPP年总量的最大值出现在2010年,为2 023.18 g·m-2·a-1;GPP年总量的最小值出现在2005年,为1 345.68 g·m-2·a-1。
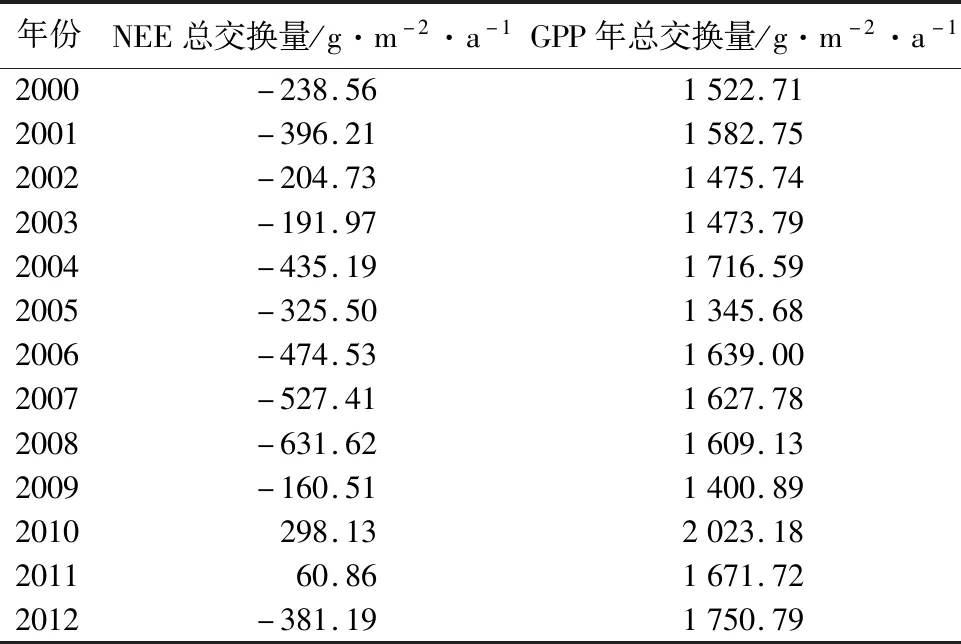
表2 2000—2012年哈佛森林生态系统NEE和GPP年总量特征
由图1可知,2000—2012年哈佛森林生态系统CO2通量与气温变化有较好的相关性(P<0.01),其中,NEE与气温的线性拟合分析的相关系数(R2)为0.65,表现为随着气温的上升,NEE有下降的趋势。GPP与气温的线性拟合分析的相关系数(R2)为0.81,表现为随气温的上升,GPP有上升的趋势。造成该现象的原因是由于气温的上升促进了哈佛森林生态系统中木本植物的生长,增强了植物群落的光合作用,造成了NEE的下降趋势以及GPP的上升趋势。
3.2 基于物候模型的CO2通量变化量计算
3.2.1 NEE变化量计算
由表3可知,将2000—2012年NEE数据经物候模型拟合后,每年拟合结果的相关系数(R2)均大于0.8,即代表将物候模型应用至该地区的NEE数据拟合的效果较好。每年的NEE变化量特征较相似但NEE变化量的最小点、最大点、0点出现的日序不同,且变化值不同。2000—2012年NEE变化量最小点所在日序范围为129~181 d。NEE变化量0点所在日序范围为169~221 d,NEE变化量最大点所在日序范围为225~294 d。其中NEE变化量最小点所出现的最早日序的年份为2010年,变化值为-0.50 g·m-2·d-1,即认为2010年在该天的NEE下降速率达到最大[17],可代表植物群落光合作用增强幅度最大的时间点。NEE变化量0点所出现的最早日序的年份为2012年,即认为2012年在该天的NEE总量达到最低,可代表植物光合作用最强的时间[17]。NEE变化量最大点所出现的最早日序的年份为2012年,变化值为0.07 g·m-2·d-1,即认为2012年在该天的NEE的上升速率达到最大。2000—2012年中NEE变化量最小点所在日序的平均值为152 d,NEE变化量0点所在日序的平均值为194 d,NEE变化量最大点所在日序的平均值为255 d,依据NEE的动态特征分析的结果,推测哈佛森林生态系统的生长季中点为第194 d[17]。NEE变化量的最小点、0点、最大点出现的变化值不同,其中2000—2012年中NEE变化量最小点的范围为-0.55~-0.11 g·m-2·d-1,平均值为-0.27 g·m-2·d-1,NEE变化量最大点的范围为0.07~0.16 g·m-2·d-1,平均值为0.11 g·m-2·d-1。基于2000—2012年NEE变化量最小点、0点、最大点的平均日序分析,推断哈佛森林生态系统的生长季光合作用最强的时间开始于春季末期,结束于秋季初期,主要集中于夏季中的3个月中。该结果与Richardson et al.[13]研究多通量站点森林生态系统物候特征较相似,与Gu et al.[17]研究芬兰温带森林生态系统的生长季中点在200 d的结果较一致。
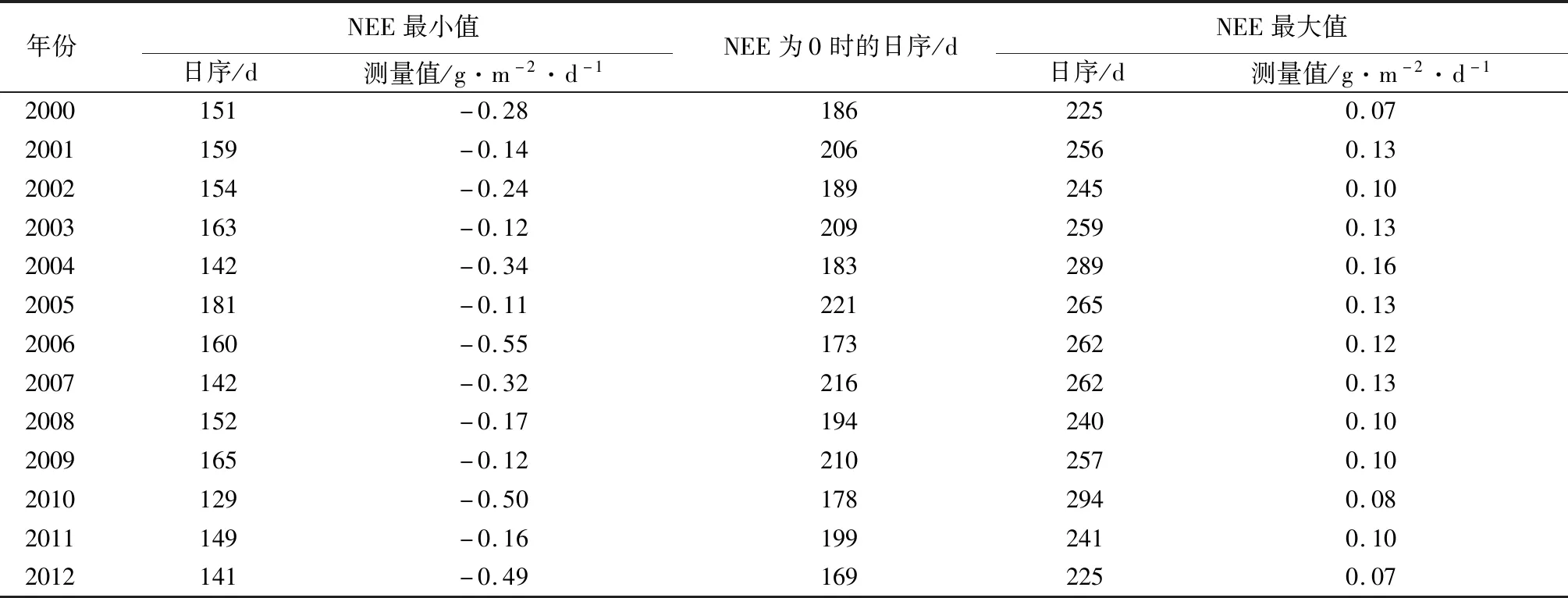
表3 2000—2012年哈佛森林生态系统NEE变化特征
3.2.2 GPP变化量计算
由表4可知,2000—2012年GPP数据经物候模型拟合后,每年拟合结果的相关系数(R2)均大于0.8,即代表将物候模型应用至该地区的GPP数据拟合的效果较好。2000—2012年中GPP变化量最大点所在日序范围为130~172 d。GPP变化量0点所在日序范围为169~209 d,GPP变化量最小点所在日序范围为215~305 d。其中GPP变化量最大点出现的最早日序的年份为2007年,变化值为0.23 g·m-2·d-1,即认为2007年在该天GPP上升速率达到最大,可代表植物光合作用增强幅度最大的时间。GPP变化量0点出现的最早日序的年份为2006年,即认为2006年在该天GPP总量达到最高,可代表植物光合作用最强的时间[17]。GPP变化量最小点出现的最早日序的年份为2008年,变化值为-0.15 g·m-2·d-1,即认为2008年在该天GPP的下降速率达到最大。2000—2012年GPP变化量最大点所在日序的平均值为150 d,GPP变化量0点所在日序的平均值为191 d,GPP变化量最小点所在日序的平均值为256 d,这与NEE变化量最小点所出现的日序的平均值为152 d,NEE变化量0点所出现的日序的平均值(194 d),NEE变化量最大点所出现的日序的平均值(255 d)的结果较相似。依据GPP动态特征的分析,推测哈佛森林生态系统植物进入光合作用稳定期的时间约开始于全年的第150 d,中点为第191 d,结束于第256 d,全年约共持续106 d。基于GPP动态特征的分析,依据北半球季节的一般划分,推断哈佛森林生态系统的生长季光合作用稳定期开始于春季末期,结束于秋季初期,且主要集中于夏季的3个月中,这与NEE动态特征的分析结果一致。
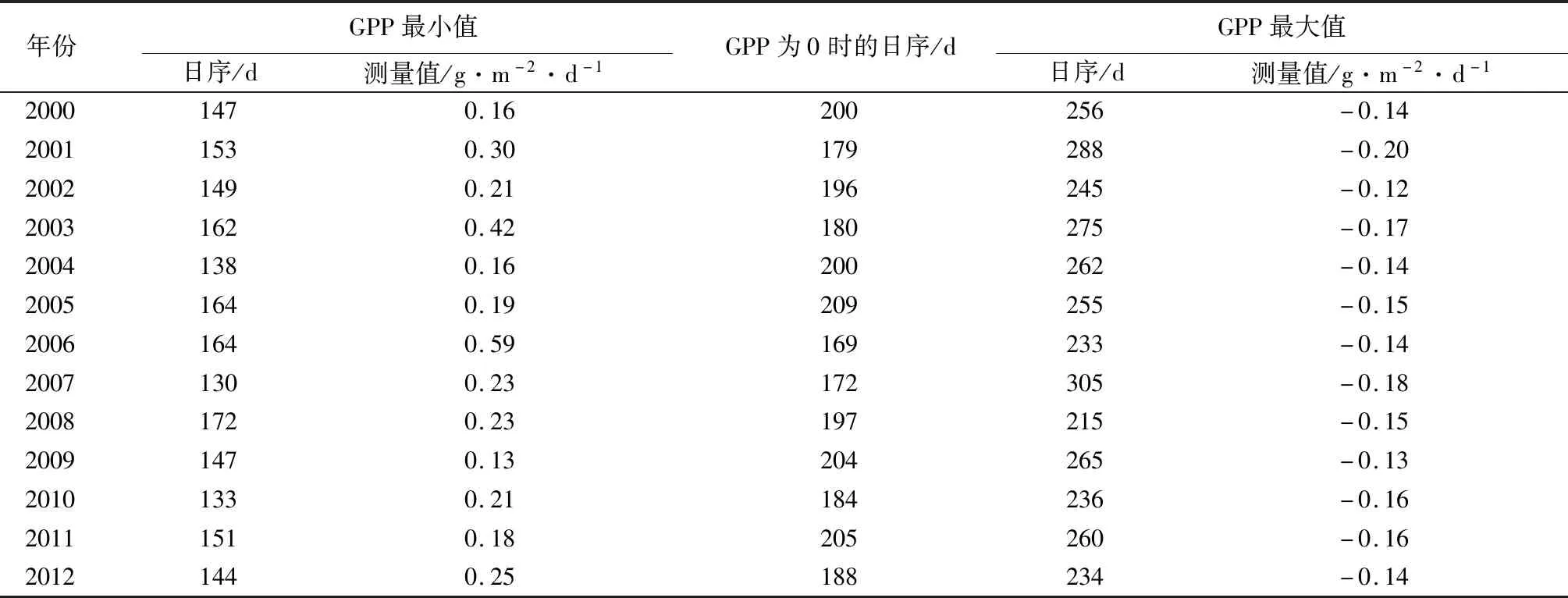
表4 2000—2012年哈佛森林生态系统GPP变化特征
3.3 CO2通量对气温变化的响应
由表5可知,2000—2012年气温数据经物候模型拟合后,每年拟合结果的相关系数(R2)均大于0.8,即代表将物候模型应用至该地区气温数据拟合的效果较好。2000—2012年气温变化量最大点所出现的日序范围为79~151 d,平均值为105 d。气温变化量0点所出现的日序范围为201~270 d,平均值为212 d。气温变化量最小点所在的日序范围为255~315 d,平均值为287 d。
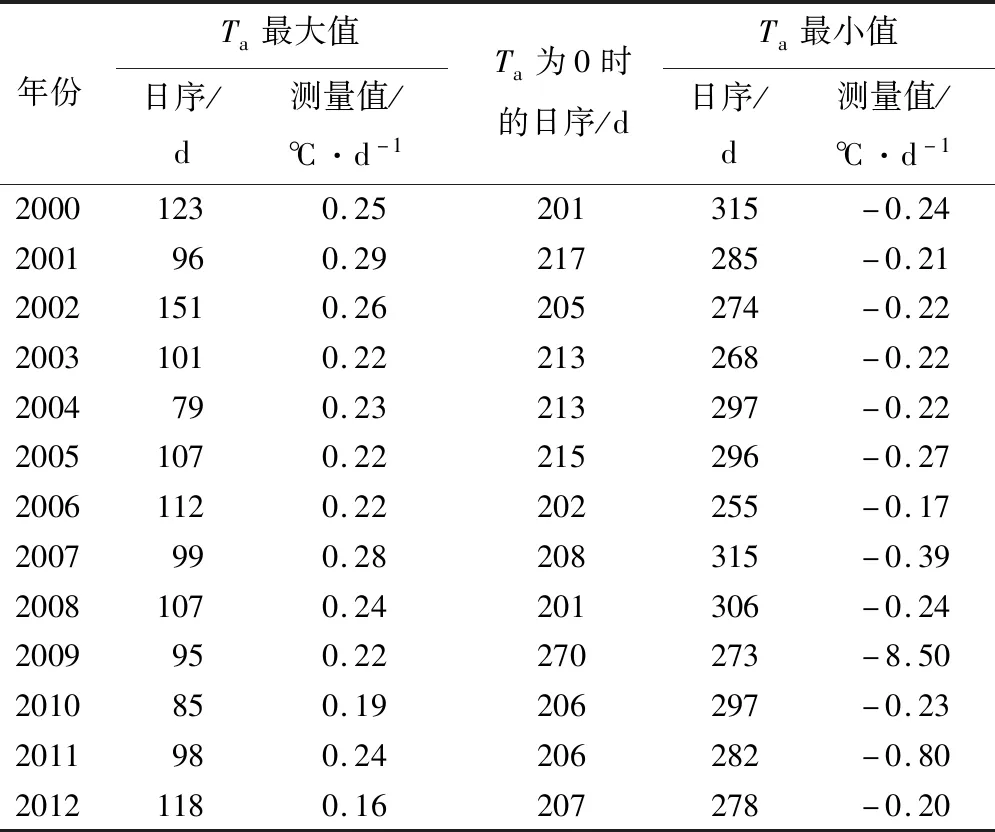
表5 2000—2012年哈佛森林生态系统气温变化特征
由表6可知,气温变化量最大点、0点、最小点所在日序,与NEE、GPP变化量最大点、0点、最小点所在日序进行拟合的相关系数(R2)均为0.81,即代表气温变化与CO2通量变化存在一定的相关性(P<0.01),且是正相关关系。即随着气温变化的时间点的推迟或者提前,会造成CO2通量变化的时间点的推迟或者提前。该分析表明气温是影响森林生态系统CO2通量的主要因素之一,与Niu et al.[15]、Richardson et al.[13]、Bracho et al.[33]研究结果一致。

表6 CO2通量对气温变化的响应
4 结论
该研究基于由国际通量网所提供的美国哈佛森林通量观测站所观测和记录的2000—2012年CO2通量数据和气温数据,使用物候模型分析了该区温带森林生态系统CO2通量的变化量特征,以及CO2通量对气温变化的响应。该森林生态系统在2010和2011年表现为碳源,在其它年份的年NEE总量均为负值,是一个较稳定的陆地生态系统碳汇;研究区植被生长状况是生态系统碳循环的主导因素,植物群落光合作用最强的时间主要集中在夏季;NEE、GPP和气温存在一定的相关性,即随着气温增长时间点的推迟或者提前,会造成CO2通量变化的时间点的推迟或者提前。
猜你喜欢
农业工程学报(2022年12期)2022-09-09
贵州大学学报(自然科学版)(2022年4期)2022-07-11
热带作物学报(2022年6期)2022-07-08
农业灾害研究(2022年1期)2022-05-07
环境(2021年5期)2021-06-20
暴雨灾害(2021年2期)2021-04-02
农业灾害研究(2020年8期)2020-05-11
Coco薇(2016年5期)2016-06-03
考试周刊(2016年6期)2016-03-11
少儿科学周刊·儿童版(2015年10期)2015-11-07
