
‘凤丹’牡丹花色变化过程中花瓣色素及相关基因表达1)
2020-05-29 00:54杨晨李厚华朱佳宾韩美玲申婷孟佳欣
东北林业大学学报 2020年5期
杨晨 李厚华 朱佳宾 韩美玲 申婷 孟佳欣
(西北农林科技大学,杨陵,712100)
牡丹(Paeoniasuffruticosa)作为中国十大传统名花之一,栽培历史悠久,有红、粉、紫、白、黄、黑、绿、蓝和复色九大色系[1]。增加花色的多样性是观赏植物的重要育种目标,牡丹的花色虽然丰富,但少有变色品种。‘凤丹’牡丹在开花过程中存在着花色变化的现象,花的颜色由紫红色逐渐变白,这增加了‘凤丹’花的观赏价值。花色的形成主要受到花色素种类和质量分数的影响[2],决定牡丹花呈色的色素是类黄酮化合物[1]。花青苷作为最重要的类黄酮化合物之一,是影响许多植物花色形成的关键性物质,其质量分数与花色变化密切相关[3-4]。
花青苷的生物合成和积累受一系列结构基因和调节基因的调控[5]。结构基因主要包括苯丙氨酸解氨酶(PAL)、查耳酮合成酶基因(CHS)、查耳酮异构酶基因(CHI)、黄烷酮羟化酶基因(F3H)、类黄酮羟化酶基因(F3’H)、二氢黄酮醇还原酶基因(DFR)和花青素合成酶基因(ANS)。调节基因(转录因子基因)在转录水平上调控结构基因的表达,与花青苷合成相关的转录因子主要包括MYB、bHLH和WD40三大类[6],它们通常以复合物的形式调节花青苷生物合成中结构基因的表达。在牡丹中,已有与花青苷合成相关的基因,如PsCHS1、PsCHI1、PsDFR1、PsMYB114L和PsMYB12L被克隆,并进行了功能验证[7-10]。
现在关于植物花色变化的研究,大多是分析了开花过程中花的色素组成及质量分数,例如,对芍药(Paeonialactiflora)、迎红杜鹃(RhododendronmucronulatumTurcz)、腊梅(ChimonanthuspraecoxLink)开花过程中花色变化的研究,普遍认为花青苷质量分数与花色变化密切相关[11-13]。Guo et al.[14]对利用转录组测序分析了芍药‘Coral Sunset’花色变化过程中与花青苷合成相关的结构基因与转录因子,总体而言,目前在分子层面上对花色变化的研究还较少。本研究以紫红色‘凤丹’花(Paeoniaostii‘feng dan’)为材料,通过对5个不同开花时期花瓣的色素成分及质量分数和花青苷合成途径中相关基因的表达水平进行分析,初步探讨了‘凤丹’牡丹开花过程中花色变化的原因,为深入研究牡丹花色变化的分子机理及花色育种提供参考。
1 材料与方法
1.1 植物材料
‘凤丹’牡丹花取自西北农林科技大学牡丹种质资源圃。选取生长健壮,长势一致,花蕾较多的植株,采集破绽期(S1)、初开期(S2)、盛开期(S3)、盛开末期(S4)、谢花期(S5)共5个开花时期的花瓣(图1)。用液氮速冻后,放入冰箱-80 ℃保存待用。
1.2 花色表型测定
使用色彩色差仪(CR-400,Japan)测量每个时期花瓣内表面除色斑外中央部分的明度L*值、色相a*和b*值。根据公式C*=(a*2+b*2)1/2计算彩度C*值。每个时期花瓣测量3朵花,每朵花测量3次。
1.3 色素提取
分别取适量不同时期新鲜花瓣,用液氮冷冻研磨成粉末,称取0.3 g粉末加入4 mL甲醇于冰箱内4 ℃遮光萃取48 h。以4 000 r·min-1离心10 min,取上层上清液,用孔径0.22 μm的过滤器过滤后用于色素测定。
1.4 色素定性定量分析
通过高效液相色谱-二极管阵列检测法(HPLC-DAD)对色素进行定性和定量分析[15]。使用日本岛津LC-2030C 3D高效液相色谱仪进行检测。设置仪器参数:柱温40 ℃,进样量10.0 μL,流速0.50 mL·min-1,190~700 nm全波长扫描吸收光谱。流动相参数:A为体积分数为0.04%的甲酸水溶液,B为色谱级乙腈。梯度洗脱程序:0 min,95%A,5%B;40 min,60%A,40%B;45 min,0%A,100%B;70 min,95%A,5%B;分析时间80 min。用甲醇溶解色素标准品,配制成500、250、100、75、50、10 mg·L-16种质量浓度,根据HPLC结果制作标准曲线。每个时期花瓣测定3次重复,样品结果通过与标准品保留时间及紫外吸收峰对比进行定性分析,质量分数根据相应的标准曲线进行计算。
1.5 qRT-PCR分析花色相关基因表达
使用Omega试剂盒提取不同发育阶段花瓣总RNA,使用PrimeScriptTMRT reagent Kit with gDNA Eraser试剂盒(TAKARA,大连)合成第一链cDNA。试验所选的转录因子根据已报道的对牡丹花青苷合成较为关键的转录因子进行选择[16]。内参基因为GAPDH,所测基因引物参考Zhang et al.[16]、Zhao et al.[17]、Gao et al.[18]设计,引物序列见表1。按照试剂盒2×Plus SYBR real-time PCR mixture(Biotake,北京)说明书,采用SYBR Green法在Applied Biosystems StepOne Plus实时定量PCR仪(Applied,USA)上进行操作。反应条件为94 ℃预变性1 min,94 ℃ 15 s,60 ℃ 15 s,72 ℃ 30 s,共40个循环。每个时期样品重复3次,采用2-△△Ct法计算基因的相对表达量。

表1 引物序列
1.6 数据分析方法
使用Microsoft Excel进行数据处理与作图,使用IBM SPSS Statistics 22.0进行方差和相关性检验。
2 结果与分析
2.1 ‘凤丹’牡丹开花过程中花色变化
‘凤丹’从破绽期到盛开期花色为紫红色,盛开之后花色逐渐变浅,到谢花期花瓣基本变白(图1)。经过花色表型测定之后发现,红绿属性a*值和花瓣彩度C*值随着花的开放呈现出逐渐下降的趋势,a*值在各个时期差异显著,C*值前两个时期差异不显著,后3个时期差异显著。从破绽期到盛开期a*和C*值降低较少,而从盛开期到谢花期降幅较大,红色减弱,彩度降低,这与‘凤丹’花从紫红色变白过程表现一致。明度值L*从破绽期到谢花期逐渐升高,即在花色变化过程中花的明度随之增强(表2)。

表2 ‘凤丹’牡丹开花过程中不同时期花色参数
注:表中数据为平均值±标准差;b*值由负到正,代表蓝色减退,黄色增强;同列不同小写字母表示Duncan’s检验在P=0.05下差异显著。
2.2 ‘凤丹’牡丹花瓣色素定性和定量分析
‘凤丹’花瓣的HPLC结果表明,花中含有芍药花素-3,5-二葡糖苷(Pn3G5G)和矢车菊素-3,5-二葡糖苷(Cy3G5G)两种花青苷(表3)。Pn3G5G和Cy3G5G的质量分数在花开放过程中呈现下降趋势,且S3到S5时期花青苷质量分数大幅减少。Pn3G5G在各个时期的质量分数占相应时期总花青苷质量分数的95%以上,远远高于Cy3G5G,是‘凤丹’花的主要花青苷。总花青苷质量分数从S1到S5时期一直下降,S5时期较S1时期质量分数降低了84.11%。
此外,还检测出了芹菜素、山奈酚、金圣草黄素、槲皮素、木犀草素、儿茶素、对香豆酸和柚皮素共8种单体酚。除金圣草黄素和对香豆酸外,其他单体酚的质量分数均在S1时期最高。芹菜素、山奈酚和柚皮素的质量分数呈先下降后上升趋势,从S1到S4时期逐渐减少,S5时期质量分数少量上升。金圣草黄素质量分数在花开放过程中逐渐升高,但是质量分数一直处于较低水平。儿茶素质量分数也较低,S1到S3时期质量分数下降,花开放后期质量分数逐渐回升。对香豆酸在盛开后质量分数下降,但5个时期的质量分数无显著差异。槲皮素、木犀草素质量分数随着花的开放而逐渐减少。芹菜素、槲皮素和柚皮素的质量分数始终高于其他单体酚(表3)。
根据HPLC结果,计算得到各个时期的总黄酮质量分数。总黄酮质量分数与花青苷质量分数变化趋势一致,呈下降趋势。S5时期总黄酮质量分数较S1时期下降了64.61%。
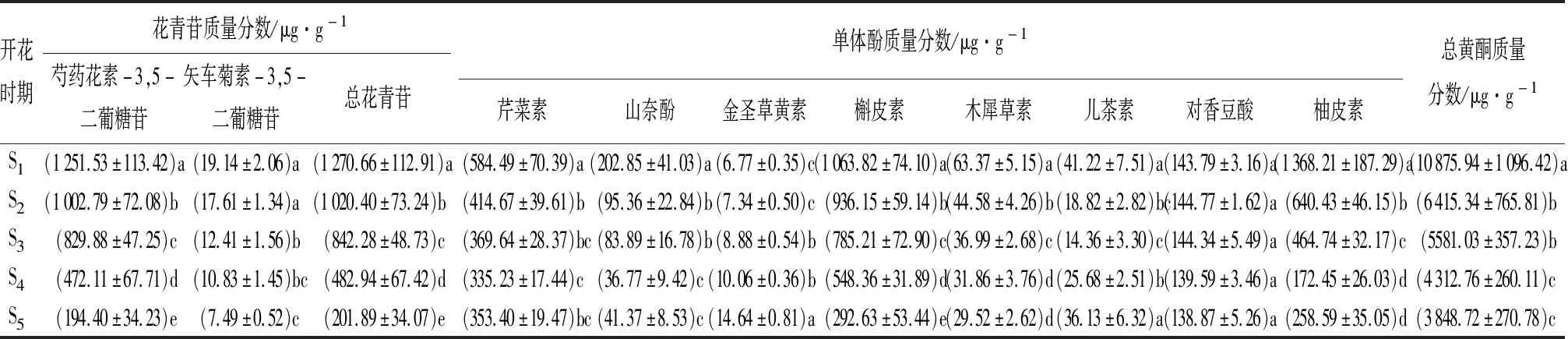
表3 ‘凤丹’牡丹不同开花时期花瓣中类黄酮成分及质量分数
注:表中数据为平均值±标准差;同列不同小写字母表示Duncan’s检验在P=0.05下差异显著。
2.3 不同开花时期花青苷合成相关结构基因的表达分析
通过荧光定量PCR对‘凤丹’5个开花时期的花青苷合成相关结构基因PAL、CHS、CHI、F3H、F3’H、FLS、DFR和ANS的表达量进行了分析。结果显示,CHS、CHI、F3’H、FLS基因的表达模式均为先上升再下降,并在S5时期表达量最低,除CHI基因外其他基因的表达量均显著低于前4个时期(表4)。CHS和CHI基因在S4时期表达量最高,而F3’H和FLS基因在S2时期表达量达到最大值。F3H基因的表达模式规律不明显,表达量先升高后下降,又升高再下降,表达量与总花青苷质量分数呈极显著负相关(图2)。PAL、DFR和ANS基因的相对表达量随着花的不断开放而逐渐降低,这与总花青苷质量分数的变化趋势一致,且呈极显著正相关(图2)。DFR和ANS在S1时期的表达量分别是S5时期的259.18、68.14倍,相比较于其他基因大量表达。推测下游结构基因DFR和ANS对‘凤丹’花青苷的合成起着重要作用。
2.4 不同开花时期花青苷合成相关调节基因的表达分析
采用qRT-PCR法测定了MYB22、bHLH1和WD40-1在不同开放时期‘凤丹’花中的表达量。WD40-1的相对表达量从S1时期到S3时期不断上升,接着逐渐降低,在S5时期质量分数最低(表4)。WD40-1与大部分结构基因在不同开花时期的表达量呈正相关(图2)。MYB22和bHLH1从S1到S5时期表达量呈下降趋势(表4),这与总花青苷质量分数,结构基因PAL、DFR、ANS的表达量变化趋势一致,且均呈极显著正相关。MYB22和bHLH1两者之间的表达量也呈极显著正相关(图2)。
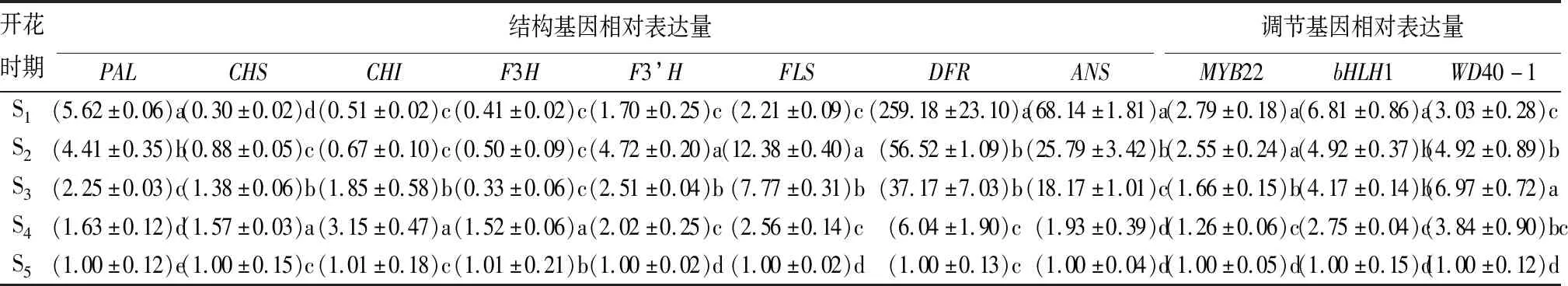
表4 ‘凤丹’牡丹不同开花时期花青苷合成相关基因的相对表达量
注:表中数据为平均值±标准差;同列不同小写字母表示Duncan’s检验在P=0.05下差异显著。
3 结论与讨论
类黄酮是高等植物花瓣呈色过程中重要的次生代谢物质,其中花青苷质量分数是影响花色的主要因素[19-22]。通过对‘凤丹’5个开花阶段的花瓣色素组成及质量分数进行分析发现,总花青苷和总黄酮质量分数降低与花色变化有关,尤其是总花青苷质量分数显著减少,是花色由紫红色向白色方向变化的主要原因。在其他植物中也有类似的结果。在鸳鸯茉莉(BrunfelsiaacuminataBenth.)中花青苷质量分数减少造成了花瓣由深紫色渐变为白色[23]。Guo et al.[14]认为两种芍药花色变化与花青苷和总黄酮质量分数有关,花青苷质量分数的减少是其从珊瑚色变为黄色的主要原因。杨琴等[15]分析了‘金衣花脸’和‘霞光’这两个牡丹品种花瓣色素,他们认为花青苷质量分数的急剧减少可能是导致花色由红向黄色变化的主要因素。
花青苷生物合成的结构基因分为早期生物合成基因(early biosynthetic genes,EBGs)和晚期的生物合成基因(late biosynthetic genes,LBGs),EBGs位于花青苷生物合成途径的上游,包括CHS,CHI,F3H等基因,负责编码黄酮、黄酮醇等类黄酮合成的酶,而LBGs包括DFR和ANS等花青苷生物合成的下游基因,主要编码合成花青苷的酶[24]。‘凤丹’花中CHS和CHI这两个基因的表达量先逐渐增加到S4,达最大值,再下降。红色芍药‘Hongyanzhenghui’花在开放过程中CHS和CHI基因呈现类似的表达模式,表达量都在盛开后达到最大,谢花期减少[26]。F3H基因表达量呈现两升三降变化趋势,与总花青苷质量分数一直下降的趋势不一致,其表达量与总花青苷质量分数呈极显著负相关,可能与F3H是花青苷合成途径上游基因,除参与花青苷合成外还参与其他的类黄酮合成有关[25,27]。DFR、ANS基因是花青苷合成的下游基因,DFR催化二氢黄酮醇形成不稳定的无色花青素,ANS将无色花青素催化成有色花青素[27]。‘凤丹’花发育过程中,花青苷合成结构基因中只有PAL、DFR、ANS基因的表达量与花青苷质量分数的变化趋势一致,且呈极显著正相关。PAL和ANS基因的表达模式与Wu等[28]的对双色芍药花的研究类似,都呈下降趋势,且与花中总黄酮质量分数减少相关。在芍药‘Coral Sunset’花色变化研究中认为PlF3’H、PlFLS、PlDFR、PlANS和PlUFGT结构基因在开花前期的高表达促进了花青苷合成,而后几个时期表达量逐渐减少导致了花青苷质量分数显著减少,造成花色变化[14]。在红色玫瑰(Rosarugosa)花中,RrDFR、RrANS高度表达导致了花青苷大量积累[29]。菊花(ChrysanthemummorifoliumRamat.)花色变化中发挥关键作用的基因包括CHS、CHI、DFR和ANS等[30]。由此推测,PAL、DFR、ANS基因对‘凤丹’花发育过程中花青苷合成起重要作用。
转录因子MYB、bHLH和WD40可通过调节花青苷生物合成途径中不同结构基因的表达水平,从而调控花青苷的合成与积累[31-33]。研究中,MYB22的表达量与基因DFR和ANS的表达量和花青苷质量分数变化趋势一致,且呈极显著正相关。近年来,对三色龙胆(Gentianatricolor)、裂叶牵牛(Ipomoeanil)等多种植物进行研究表明,MYB是植物花青苷合成中的关键转录因子[34-35]。苹果(Mulusdomestica)中MdMYB1可以正向调控果皮中MdDFR和MdANS的表达及花青苷的生物合成与积累[36]。葡萄(Vitisvinifera)中的VvMYB5a和VvMYB5b可以激活VvANS的启动子从而调控浆果的花青苷合成与积累[37]。‘岛锦’牡丹(P.suffruticosa‘Shima Nishiki’)花发育的5个时期中转录因子PsMYB12L与DFR、ANS基因的表达模式相似,PsMYB12L上调这两个基因的表达使花青苷质量分数增加[10]。因此,推测MYB22可能是‘凤丹’花青苷合成与积累中重要的转录因子。
转录因子bHLH最重要的功能之一就是调节花青苷合成[38]。在矮牵牛(Petuniahybrida)中bHLH基因Ahl可以调节PhDFRA的表达[39]。金鱼草(Antirrhinummajus)中的Delila基因(bHLH基因)能促进花青苷的合成[40]。具有与bHLH相互作用基序的MYB基因,可以形成MYB-bHLH复合物来调节花青苷生物合成结构基因的表达[41]。石斛兰(Dendrobiumhybrids)中DhMYB2与DhbHLH1相互作用以正向调控石斛兰花瓣中的花素苷产生,影响花色[42]。菊花CmbHLH2与CmMYB6转录因子共表达时显著上调CmDFR的表达,并造成花青苷的积累[43]。‘凤丹’花中bHLH1的表达模式不仅与花青苷的变化趋势,MYB22、DFR和ANS基因的表达模式一致,而且MYB22和bHLH1的表达量具有极显著相关性。由此推测,bHLH1对‘凤丹’花青苷合成起重要的调控作用,而且可能以MYB-bHLH复合物形式调控结构基因的表达,从而影响‘凤丹’花青苷的合成与积累。
综上所述,‘凤丹’牡丹花由紫红色向白色变化主要是由于总花青苷质量分数的大量减少。推测开花过程中花青苷合成关键结构基因PAL、DFR、ANS表达量的降低,使得‘凤丹’牡丹花青苷的生物合成与积累减少,MYB22和bHLH1可能主要调控结构基因DFR和ANS的表达,但具体的调控机制还需进一步研究。
猜你喜欢
课堂内外(高中版)(2021年7期)2021-01-17
初中生学习指导·提升版(2020年10期)2020-09-10
支部建设(2019年24期)2019-09-28
金桥(2019年12期)2019-08-13
文苑(2019年14期)2019-08-09
新作文·小学高年级版(2017年10期)2017-11-13
红蜻蜓(2017年8期)2017-10-30
学苑创造·A版(2016年10期)2016-11-19
小天使·四年级语数英综合(2014年4期)2014-04-04
宠物世界·猫迷(2013年2期)2013-04-27
