
流行性乙型脑炎病毒NS4B蛋白的原核表达及其结构分析
2020-05-28 09:57童剑军何川川米丽开姆托合提尼亚孜张雪萍段彦祥李有文
塔里木大学学报 2020年1期
童剑军 何川川 米丽开姆·托合提尼亚孜 张雪萍 段彦祥 李有文,2*
(1 塔里木大学动物科学学院,新疆 阿拉尔843300)
(2 新疆生产建设兵团塔里木畜牧科技重点实验室,新疆 阿拉尔843300)
(3 塔里木大学生命科学学院,新疆 阿拉尔843300)
流行性乙型脑炎(Epidemic encephalitis type B),简称乙脑,于1934年在日本发现,因此又被称为日本乙型脑炎(Japanese encephalitis)。该病作为一种人兽共患传染病,早已引起了业内外人士的广泛关注。本病主要流行于东南亚等地区,典型症状表现为意识障碍、体温升高,昏迷,更甚者可出现神经性症状[1,2],在家畜中尤以猪感染最为严重[3,4]。主要表现为突然发病,体温呈稽留状态。妊娠期母猪出现高热、流产、死胎等症状,公猪则出现生殖障碍[5,6],严重影响了我国畜牧产业的健康发展。世界卫生组织也将其列为重点防控对象[7]。
乙型脑炎病毒作为黄病毒科的一种单链RNA病毒,由11 000 个核苷酸所构成,含有一个特殊结构---多聚蛋白编码框。该框能够编码三种结构蛋白(C、M、E)以及七种非结构蛋白(NS1、NS2A、NS2B、NS3、NS4A、NS4B、NS5)[8],而其中的NS4B 蛋白在病毒复制过程中发挥重要作用[2]。据有关资料显示,黄病毒科病毒由膜相关的复合体介导复制,ns4b 与ns2a、ns2b 和ns4a 三种基因共同构成该复合体[9]。但其他黄病毒科病毒的研究结果显示,ns4b与内质网相关性更高,然而其相关机制尚不明确。Tajima 等人认为黄病毒科中NS4A 与NS4B 发生互作[10],但Yu 等却发现在Vero 细胞中,西尼罗河病毒(West Nile Virus)的NS4A 与NS4B 未互作[11]。由此可知在整个黄病毒科中,不同病毒种的非结构蛋白作用方式差异显著。NS4B 蛋白自身就是一个典型的跨膜蛋白,有三个跨膜区域,来回穿梭于膜中,其可能对膜的活性、膜的物质和能量交换意义重大。在其致病过程中,病毒自身的蛋白质功能起着关键的作用,但我们对JEV 的NS4B 蛋白的功能知之甚少,为研究NS4B 蛋白的功能并进一步揭示其分子作用机制,本研究拟获得乙型脑炎P3 株ns4b 基因的重组表达蛋白,预测蛋白性质和功能,以期进一步研究该蛋白的功能。
1 材料
1.1 菌种与载体
乙脑病毒P3 株由华中农业大学曹胜波教授馈赠;大肠杆菌DH5α、BL21 以及原核表达载体均保存于新疆生产建设兵团塔里木畜牧科技重点实验室。
1.2 主要药品及试剂
Primestar HS DNA Polymerase、dNTP、T4 DNA 连接酶(购自大连宝生物科技有限公司),琼脂糖凝胶回收试剂盒(购买北京天根生物工程技术服务有限公司),Nde I、Xho I(购自Thermo 公司),DNA 纯化试剂盒(购自美国Omega 公司)。其他试剂按说明书配制。
1.3 主要使用仪器
DYY-12 琼脂糖水平电泳仪(北京六一生物科技有限公司),全自动凝胶成像分析仪、HC 高电流电泳仪、T100Thermal Cycler 梯度PCR 仪、Mini-PROTEAN Tetra 电泳槽、Mini-Trans-Blot 电泳转印槽(Bio-Rad生命科学有限公司),XMTP-204 梯三孔三温水浴锅(上海博讯实业有限公司医疗设备厂),SWCJ-2FD 超净工作台(BXOXUN)等。
2 方法
2.1 原核表达载体的构建
2.1.1 ns4b基因特异性引物设计
参考GenBank 中公布的乙型脑炎病毒JaOArS982 参考株(登录号:NC001437)4B 基因序列,用DNAstar 软件设计特异性引物,送公司合成。如表1(下划线为加入酶切位点:Nde I/Xho I)。

表1 引物合成表
2.1.2 ns4b基因的PCR扩增与产物回收
以乙型脑炎病毒P3 株基因组cDNA 为模板,进行PCR 扩增。PCR 产物纯化回收并鉴定(按试剂盒说明书操作)。
2.1.3 原核表达载体pET-42b-NS4B的构建及鉴定
用Nde I 和Xho I 双酶切pET-42b 质粒和回收的PCR 产物,酶切后产物利用DNA 纯化试剂盒纯化并收集。将二者连接后转入大肠杆菌感受态DH5α,涂板培养筛选获得阳性菌提取质粒,并用双酶切(Nde I/Xho I)鉴定,质粒送往生工生物工程(上海)股份有限公司测序鉴定。
2.2 NS4B蛋白表达鉴定
将阳性克隆质粒转化至感受态细胞BL21,对其表达产物进行SDS-PAGE 电泳,并进行Western Blot检测,具体参照李有文[12]的方法。
2.3 NS4B蛋白的生物信息学分析
2.3.1 蛋白信号肽及跨膜结构的预测
NS4B 蛋白信号肽的预测,使用在线软件服务器SignalP4. 1Server,网址为http://www. cbs. dtu. dk/services/SignalP/;NS4B 蛋白跨膜结构的预测,使用在线软件服务器TMHMMServerv. 2. 0,网址为http://www.cbs.dtu.dk/services/TMHMM/。
2.3.2 蛋白糖基化位点、磷酸化位点和抗原决定簇的预测
NS4B 蛋白糖基化位点和磷酸化位点的分析预测,使用在线软件服务器NetOGlyc4. 0Server(http://www. cbs. dtu. dk/services/NetOGlyc/)和NetPhos3.1Server(http://www. cbs. dtu. dk/services/NetPhos/)。NS4B 蛋白质抗原决定簇的预测,使用在线软件服务器PredictingAntigenicPeptides,网 址 为http://imed.med.ucm.es/Tools/antigenic.pl。
2.3.3蛋白质抗原指数和β-turn的预测
NS4B 蛋白抗原指数和β-turn 的预测,使用在线软件服务器IEDB,预测的网址为http://www. iedb.org。
2.3.4蛋白质亲水性和柔韧性的预测
NS4B 蛋白亲(疏)水性和柔韧性的预测,使用在线软件服务器ProtScale 和IEDB,预测的网址分别为http://web. expasy. org / protscale/和http://www. iedb.org。
2.3.5蛋白二级结构和三级结构的分析预测
对于NS4B 蛋白二级结构分析及预测,使用在线软件服务器SOPMA(https://prabi.ibcp.fr/htm/site/web/home)和GalaxyTBM(http://galaxy. seoklab. org/);对 于蛋白质三级结构的分析预测及立体模型构建,使用在线软件服务器Phyre2,网址为http://www. sbg. bio.ic.ac.uk/phyre2/html/page.cgi?id=index。
3 结果与分析
3.1 ns4b基因的扩增
以JEV P3株的基因组为模板PCR 扩增的ns4b基因经琼脂糖凝胶电泳检测的结果(图1),在近800 bp处有一条带,与预期大小相符。
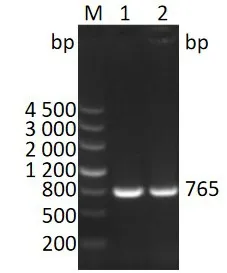
图1 ns4b基因PCR产物琼脂糖凝胶电泳
3.2 NS4B蛋白原核表达载体鉴定
3.2.1 酶切鉴定
经菌液PCR 鉴定,将阳性菌提取质粒并Nde I/Xho I 的双酶切鉴定,结果得到了与理论相符的载体和目的基因两条带(图2),鉴定为阳性克隆。
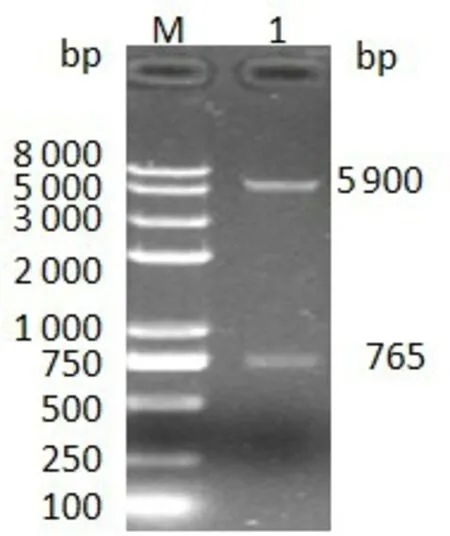
图2 pET-42b-NS4B质粒双酶切(Nde IXho I)
3.2.2 测序结果
将鉴定正确的单菌落37 ℃过夜培养,提取重组质粒pET-42b-NS4B,送往生工生物工程(上海)股份有限公司测序鉴定,结果表明P3 株NS4B 基因全长765 bp,与参考株同源性99%以上,编码255 个氨基酸,测序结果见表2。

表2 ns4b核苷酸和氨基酸序列
3.3 ns4b遗传进化序列分析
根据P3 株测序结果与GenBank 中发表的JEVns4b 的序列进行遗传进化分析(图3),可见ns4b 基因已发现的25 个毒株的遗传距离很小,只有0.8,说明其亲缘关系较近,其中P3 株该基因与JEV strain HW和JEV isolate JEV-hubei 亲缘关系最近,说明ns4b 基因非常保守,对病毒生长繁殖作用重大。
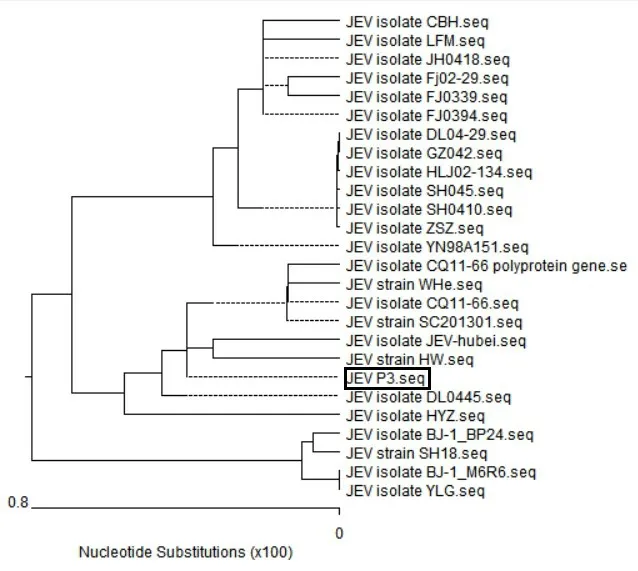
图3 ns4b遗传进化序列分析
3.4 NS4B蛋白的表达
3.4.1 SDS-PAGE 鉴定
将正确克隆的质粒转化BL21 并诱导表达的蛋白,经SDS-PAGE凝胶电泳检测。结果显示:NS4B蛋白与His 标签融合表达了27 KDa 的蛋白(图4),以包涵体形式表达在沉淀中,上清中含量很低。
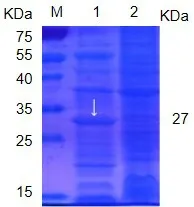
图4 pET-42b-NS4B蛋白表达
3.4.2 Western Blot检测
以His 单抗进行Western blot 检测其His 融合蛋白的表达(图5),于25 KDa-35 KDa 处出现蛋白印迹,说明融合表达成功。

图5 Western blot结果
3.5 NS4B蛋白生物信息学分析结果
3.5.1 信号肽与跨膜结构的预测结果
NS4B 无信号肽(图6)。NS4B 为膜蛋白,有255个氨基酸,1-90aa 表示在膜内区;91-113aa 表示跨膜区;113-251aa表示膜外区,有138个氨基酸残基在膜外区(图7),说明NS4B确实是一个跨膜蛋白。
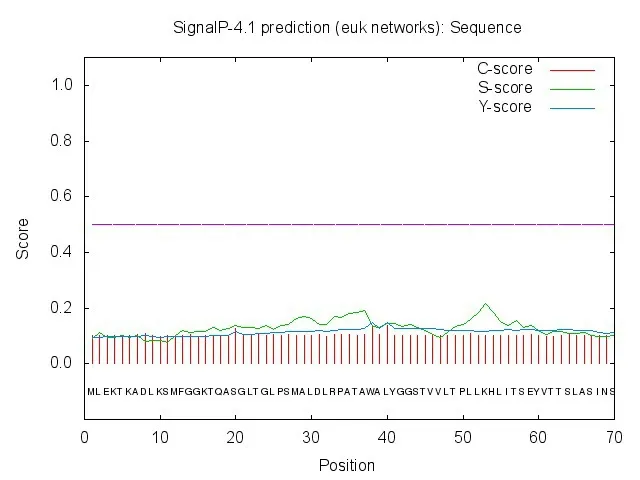
图6 NS4B蛋白信号肽预测
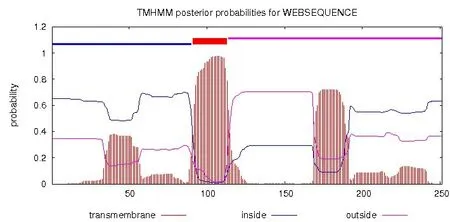
图7 NS4B蛋白跨膜结构预测
3.5.2 糖基化位点、磷酸化位点和抗原决定簇的预测结果
NS4B 蛋白共有26 个磷酸化位点(第11、20、23、44、45、49、58、60、64、67、70、74、103、137、153、162、178、190、202、204、211、216、220、229、239、249 位氨基酸)(图8)。NS4B 蛋白拥有10 个抗原决定簇,第26~34 位 氨 基 酸(PSMALDLRP)、第36~68 位 氨 基 酸(TAWALYGGSTVVLTPLLKHLITSEYVTTSLASI)、 第71~106位氨基酸(QAGSLFVLPRGVPFTDLDLTVGLVFLGCWGQITLTT)、第108~120 位 氨 基 酸(LTAMVLATLHYGY)、第126~132 位氨基酸(QAEALRA)和第142~149 位氨基酸(MKNAVVDG)、第151~157 位氨基酸(VATDVPE)、第168~190 位 氨 基 酸(KVGQVLLIGVSVAAFLVNPNVTT)、第192~203 位氨基酸(REAGVLVTAATL)、第219~233 位氨基酸(ATGLCHVMRGSYLAG)(图9)。
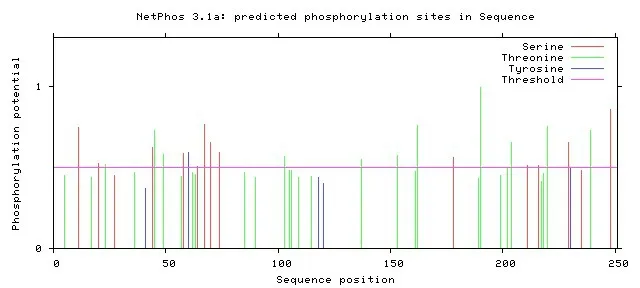
图8 NS4B蛋白磷酸化位点的预测
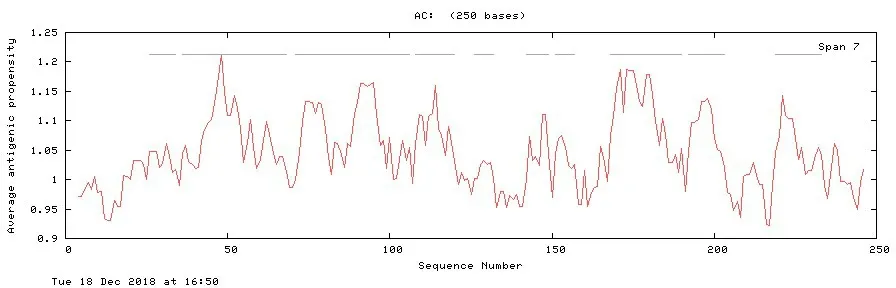
图9 NS4B蛋白抗原决定簇的预测
3.5.3 蛋白质抗原指数和β-turn的预测结果
NS4B 蛋白抗原指数最高为1.213(图10);NS4B蛋白β-turn结构最高分值为1.236,主要分布于242~248位氨基酸(图11)。
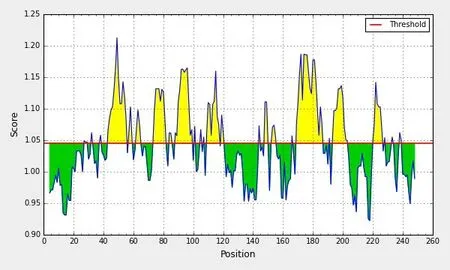
图10 NS4B蛋白抗原指数分析结果
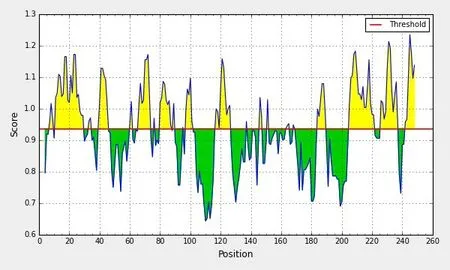
图11 β-turn 预测结果
3.5.4蛋白质亲水性和柔韧性的预测结果
分值高的氨基酸为第14~21 位氨基酸,第207~245 位氨基酸,得分最高为5. 586(图12);分值高的氨基酸区段主要为第13~21 位氨基酸、第40~46 位氨基酸、第67~76位氨基酸、第214~221氨基酸、第242~250氨基酸,分值最高为1.069(图13)。
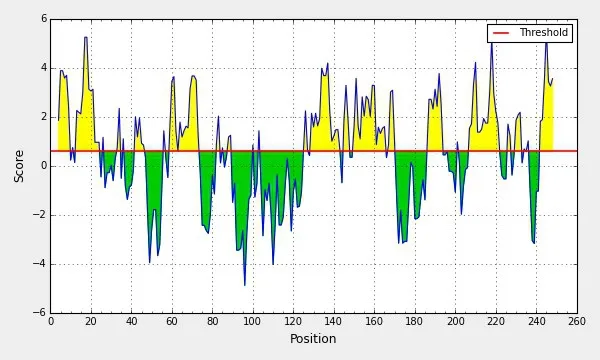
图12 NS4B蛋白亲水性预测结果
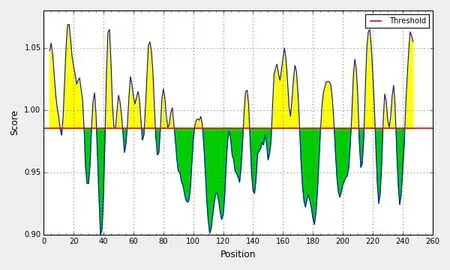
图13 NS4B蛋白柔韧性的预测结果
3.5.5 蛋白二级结构和三级结构的预测结果
NS4B 蛋白中共有129 个氨基酸构成α 螺旋,占总氨基酸数的51. 39%;有35 个氨基酸构成了延伸链,占总氨基酸数的13.94%;有17个氨基酸构成了β转角,占总氨基酸数的6.77%;有70个氨基酸构成无规则卷曲,占总氨基酸数的27.89%(图14)。在线软件服务器Phyre2 预测构建了NS4B 蛋白的三级结构模型(图15)。
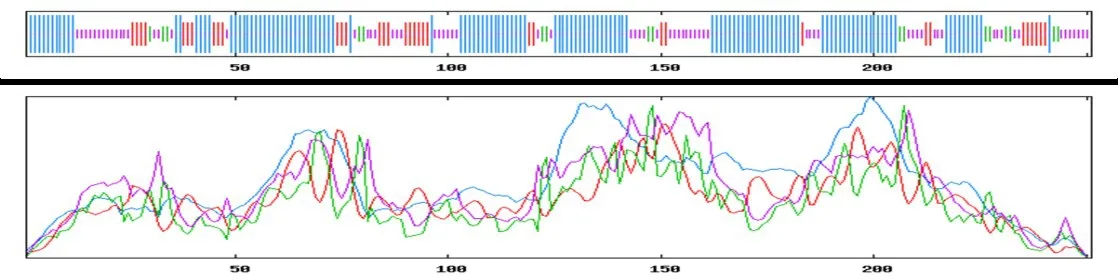
图14 NS4B 蛋白二级结构的预测

图15 NS4B 蛋白三级结构的预测
4 讨论
JEV 属于黄病毒科黄病毒属,是一种重要的神经性疾病病原,一直为人们所重视。由于黄病毒属多聚蛋白编码框的存在,使很多蛋白功能较为复杂。据研究,黄病毒属的非结构蛋白NS4B 不仅参与病毒的组装与复制,而且具有潜在的抗病毒靶点作用;另外,NS4B 可与另一个跨膜蛋白NS4A 通过2 KDa大小的信号肽连接形成异源二聚体而诱导膜重排[13];还有,NS4B 蛋白能通过诱导宿主细胞的线粒体变性抑制先天性免疫反应[14]。
本研究扩增了JEV 的P3 株ns4b 基因,其系统进化树结果显示,JEV 虽为RNA 病毒,但其NS4B 基因的保守性较强,与目前基因库中登录的25 株病毒该基因的核酸序列同源性为98. 8%以上,遗传距离仅0.8,说明NS4B 蛋白组成相当稳定,可能对病毒生长繁殖发挥重要作用。这与黄病毒属NS4B 蛋白参与病毒的组装与复制的重要功能相一致。JEV-NS4B含有26 个磷酸化位点,是一个磷蛋白。磷酸化能介导蛋白活性[15],增强蛋白相互作用的能力,因此推测JEV-NS4B 蛋白是一个活性很强的蛋白。现研究结果证明,NS4B 蛋白不仅可以自身相互作用形成同源二聚体,还可以与NS4A 蛋白通过2 KDa 信号肽连接形成异源二聚体,还可以与NS3 解旋酶相互作用[13]。这些蛋白间的相互作用是病毒复制复合体形成的分子基础,对病毒的复制至关重要[11,16,17],特别是NS4B蛋白与病毒的其他蛋白互作形成二聚体可以形成病毒复制泡,促进病毒增殖。但是对丙肝病毒(HCV)NS4B 蛋白研究发现其能通过其膜外区以及C 端的氨基酸发生自身互作,形成同源二聚体,破坏HCV NS4B二聚体的形成,能够影响复制泡的形成,抑制病毒复制[18-20]。从这个角度考虑有望利用NS4B蛋白的自聚合来抑制病毒增殖而控制疫病。
经预测JEV-NS4B 蛋白具有3 个跨膜区域,是一个标准的跨膜蛋白,原核表达得到不溶性的大小约为27 KDa 的蛋白,与其他黄病毒属该蛋白疏水性非结构膜蛋白的特性一致[8]。膜蛋白的功能之一就是作为转运体,转运体是细胞内外物质转运的分子基础,包括离子转运体、神经递质转运体、营养物质转运体以及外来物质转运体。有些药物可通过对某种转运体的抑制作用而产生效应,因此转运体可以作为药物靶标用于疾病治疗。NS4B蛋白就可作为治疗黄病毒感染的一个新型药物靶标[21]。研究表明,化合物NITD-618 能够靶向登革热病毒(DENV)的NS4B 蛋白,对DENV 具有明显的抑制效果,其EC50在低至1.0µmol,但该化合物对WNV、YFV等其他黄病毒无效[22]。马尼地平的病毒靶点是位于JEV 非结构蛋白NS4B 的130 位谷氨酰胺(Q),Q130 突变成正电氨基酸精氨酸(R)或赖氨酸(K),突变病毒对马尼地平不敏感[23]。化合物CCG-3394和CCG-4088也能靶向NS4B 蛋白,抑制黄热病毒(YFV)的复制[24]。因此加紧对NS4B 蛋白生物学特性和免疫特性的研究,才能更好地利用NS4B 靶标,开发更多更好的药物治疗黄病毒感染。
信号肽多由15~30个氨基酸组成,对蛋白质跨膜转移(定位)的N-末端的氨基酸序列有重要指导意义,在蛋白成熟后信号肽自然脱落,保证蛋白基本功能[25]。对JEV-NS4B 蛋白的氨基酸序列进行生物信息学分析发现其不含有信号肽,但是有研究表明黄病毒的NS4B 可与另一个跨膜蛋白NS4A 通过2 KDa大小的信号肽连接形成异源二聚体而诱导膜重排[13],利于病毒的复制。资料显示JEV-NS4A蛋白也不含有信号肽,这个二聚体的形成机制还有必要进一步研究。有关文献报道,信号肽结构有利于提高蛋白质的可溶性[26],无信号肽结构的蛋白易因可溶性低而更易形成包涵体。这也提示如果要得到高质量大量表达的该蛋白,在构建时加一个信号肽可能会有较好的效果。NS4B 蛋白拥有10 个抗原决定簇,且含有17 个氨基酸组成可被称为环(loop 区)的β 转角,该区为功能性抗体的识别位点的可能性很大[27],由此推测NS4B 蛋白具有功能性抗原的潜质,可作为未来抗体的备选作用靶点,也能为乙型脑炎疫苗的研发提供多重选择。NS4B 蛋白的柔韧性较为优良,二级结构内α-螺旋结构占比高达51.39%,其稳定性可见一斑,可为筛选蛋白环节提供便利,提高抗体生产效率,加快疫苗研发进程;β 折叠和无规则卷曲区域的二级结构较为松散,更易于盘旋和扭曲,该区域可能存在优势抗原表位[28],更容易与抗体分子结合,可有效促使机体产生免疫反应[29],提高其抗病力,缩短病程。本试验通过对乙型脑炎病毒NS4B 蛋白生物学信息分析,为后期更深入研究乙型脑炎病毒并发掘其潜在功能提供参考。
猜你喜欢
中国畜禽种业(2022年3期)2022-11-22
分子催化(2022年1期)2022-11-02
传染病信息(2022年3期)2022-07-15
交通医学(2022年2期)2022-06-14
烟草科技(2021年6期)2021-06-24
昆明医科大学学报(2021年1期)2021-02-07
康颐(2020年2期)2020-09-10
生物学教学(2018年4期)2018-11-29
生物学教学(2018年10期)2018-11-29
电脑知识与技术(2018年19期)2018-11-01
