
植物锌指蛋白转录因子家族研究进展①
2020-05-22 12:48:36潘介春张树伟王金英颖李浩然徐炯志彭宏祥何新华广西大学农学院广西南宁530004
热带农业科学 2020年2期
李 琳 丁 峰 潘介春 张树伟 黄 幸 王金英 王 颖李浩然 徐炯志 彭宏祥 何新华(1 广西大学农学院 广西南宁530004;
2 广西作物遗传改良生物技术重点开放实验室 广西南宁530007;
3 广西壮族自治区农业科学院园艺研究所 广西南宁530007;4 亚热带农业生物资源保护与利用国家重点实验室 广西南宁530004)
锌指蛋白存在于真核生物中,是最重要的转 录因子之一。转录因子可直接结合或间接作用于基因启动子,进而形成具有RNA聚合酶活性的动态转录复合体的蛋白质因子,通过激活或抑制基因的转录,起到调控基因表达和生物体功能等作用,完成生物在转录水平上的调控[1]。根据其功能方面的差异,转录因子可分为通用转录因子、序列特异性转录因子和辅助转录因子等。与RNA聚合酶I、II 和III 相对应的转录因子有3 类,分别是TFI、TFII 和TFIII,锌指蛋白为TFIII 型转录因子[2],它是目前发现的种类最广泛,在真核生物的生长发育以及抗逆过程中具有重要调控作用的一类核酸结合蛋白。
锌在植物生长过程中发挥着不可或缺的作用,对于锌指蛋白,锌的存在也极其重要,不仅是锌指蛋白发挥作用的关键,锌指功能的有效性也依赖于稳定的锌指结构。如果使用锌离子螯合剂脱锌或用Fe、Al、Cu、Ag、Mn 等其它金属离子代替锌离子后,锌指蛋白与DNA 或RNA 等结合特异性就会被明显抑制,不能形成折叠结构的同时,稳定的蛋白也会失去支撑力,影响基因表达及胁迫响应,使其丧失大部分功能。锌指序列主要存在于转录因子中,并在转录水平上响应及参与真核生物生长发育,并对生物及非生物胁迫做出反应[2]。
1 锌指蛋白转录因子概述
Bogenhagen等[3]于1982年发现TFIIIA参与非洲爪蟾卵母细胞5SRNA基因的调控,现已在各种植物、动物及酵母中均发现了参与基因调控的锌指蛋白[4-5],该蛋白质由一致序列的九次不完全重复组 成:(Tyr,Phe) -X-Cys-X2-4-Cys-X3-Phe -X5-Leu-X2-His-X3-4-His-X2-6(其中X 是任何氨基酸),这些阵列结合5S RNA 基因中的43 bp 序列[6],显著特征是具有“手指状”结构域且能与Zn2+结合。SPl属于典型的TFIIIA类型的锌指蛋白,其中两个Cys 和两个His 与锌离子形成配位键,从而形成一个包含β发夹和α螺旋的紧密指状结构。植物锌指蛋白中一般包含有1 到4 个数目不等的锌指结构(图1)[4]。
研究发现,拟南芥中包含176个锌指蛋白,构成了该植物中最丰富的转录因子家族,其中有33个为在其它真核生物中保守的拟南芥锌指蛋白,剩下大多数(约81%)为植物特异性锌指蛋白[2]。目前研究较为广泛的一类为真核生物基因组中含量最多的C2H2型锌指蛋白,除含有DNA 结合区DBD(DNA-binding domain)外,通常还具有参与亚细胞定位的核定位信号区NLS (nuclear localiza‐tion signal,也称B-box)[7],该蛋白与DNA 接触面有一段高度保守的氨基酸序列QALGGH,为植物锌指蛋白所特有,其中Q 若突变则会极大降低蛋白结合DNA 的能力,除Q 外其他任何一个氨基酸的突变都会导致该蛋白完全丧失结合DNA 的功能[8]。C2H2型锌指蛋白常见于植物中,不仅是染色质结构的重要组成部分,在转录调控中也发挥重要作用。
2 锌指蛋白转录因子的分类
根据锌指蛋白在指状二级结构中与锌离子结合的半胱氨酸和组氨酸残基的数量和顺序的不同,Berg 和Shi 将锌指蛋白分为9 大类:C2H2、C8、C6、C3HC4、C2HC、C2HC5、C4、C4HC3和CCCH(C 和H 分别代表半胱氨酸和组氨酸)(表1)[9];根据锌指Cys和His 残基所构成的空间结构不同,Krishna 等[10]把锌指蛋白分成8个不同的折叠群:类C2H2型锌指、高音谱号锌指、带状锌指、塞结状锌指、Zn2/Cys6 型锌指、类TAZ2 型锌指、锌离子结合短环锌指、金属硫蛋白锌指,其中前3类组成了大部分的锌指[10];根据锌指蛋白的功能差异,可分为TFIII蛋白、WRKY 蛋 白、RING 蛋白、GATA 蛋白、PHD 蛋白、DOF蛋白等亚家族。

表1 9类锌指蛋白的序列及其代表蛋白质
3 锌指蛋白转录因子家族的生物学功能
在植物生长过程中,各种不同的胁迫因素严重影响植物的生长发育及作物产量。植物在漫长的进化过程中,通过调节细胞内基因表达的模式形成了一系列的胁迫响应保护机制,越来越多的基因和蛋白在应答胁迫中发挥重要作用。其中锌指蛋白已被证实与生物体内的各种RNA 代谢有关[11],广泛参与基因的转录、翻译、mRNA 的运输、细胞支撑、蛋白折叠和染色质修饰等过程,对植物基因表达及调控起着重要作用[12]。
3.1 对植物的生长发育调控
锌指蛋白广泛参与植物的生长发育过程,例如种子萌发及成熟、根系发育、花器官及配子发育、激素反应、叶片衰老和细胞凋亡等[6]。
3.1.1 参与植物种子萌发及成熟
种子发芽受外界环境条件和内源激素的控制,ZFP3是C2H2型锌指蛋白家族的一员,在拟南芥中充当ABA,作为抑制种子发芽的负调节剂。研究表明过表达ZFP3 和与之密切相关的ZFP1、ZFP4、ZFP6及ZFP7 锌指蛋白因子使ABA 对种子发芽不敏感,而ZFP3、ZFP4 双突变体则显示出更高的ABA 敏感性。通过观察ZFP3 过表达植物的生长状况,显示了多种表型改变,例如半矮生生长习性、繁殖力缺陷以及下胚轴伸长对红光或蓝色光的敏感性增强。用光敏色素和遗传相互作用分析ABI 突变体,表现出ZFP3 通过除光敏色素A 之外的感光增强,这些数据基本可以得出:ZFP3 和锌指因子相关的ZFP 亚家族在植物发芽和幼苗早期发育期间调节光和ABA响应,影响植物的生长发育和繁殖能力,并调节幼苗光形态发生过程中的红光信号,此外ABA还控制植物对干旱和盐胁迫的响应,包括气孔关闭、与渗透调节有关的基因激活[13]。拟南芥中ARS1 编码具有一个锌指结构域、一个核输出信号(NES)域和两个核定位信号(NLS)域的核蛋白,ARS1 突变体表现出降低超氧化物歧化酶基因的表达,并在ABA处理后提高了活性氧(ROS)的积累。ARS1在拟南芥原生质体中的瞬时表达强烈抑制ABA介导的ROS产生,说明ARS1能够响应ABA和氧化胁迫,进而调节种子的萌发[13]。OsZF77基因是属于CCCH型锌指类的一个核酸结合蛋白,编码280个氨基酸,通过将OsZF77启动子区和GUS 融合基因导入水稻中观察发现:在胚乳尤其是靠近种皮外围的胚乳中GUS 基因活力较强,胚中未检测到GUS 活性。通过对OsZF77进行“RT-PCR”分析、生物信息学分析和GUS的融合表达分析表明,该基因是一个胚乳优势表达基因,结合该基因在种子中的表达模式,进一步验证了OsZF77基因的启动子在种子胚乳中的优势表达活性,也表明了该基因可能在种子胚乳发育过程中起着重要的调节作用[14]。夏苗[15]通过酵母激活实验分析证明了ZmZFP1(Zea maysL.ZFP1)具有转录激活活性,ZmZFP1转录因子具有典型的锌指蛋白二级结构、三级结构,与其亲缘关系较近的基因都含有保守结构域,通过Plant-CARE 程序预测ZmZFP1的启动子元件,发现启动子中含有ABA诱导的顺式作用元件,实时定量PCR分析得到该转录因子在胚乳中的表达结果与测序分析的表达结果一致,ZmZFP1基因上调表达可被ABA 诱导。通过检测GUS 和LUC 酶的活性也验证了ZmZFP1能提高启动子Sh2、Bt1、Wx 活性,从而使ZmZFP1能参与以及促进淀粉合成酶相关基因的表达。
3.1.2 参与植物花器官及配子发育
授粉是植物花粉和柱头、花柱传递组织之间的一系列复杂的细胞识别和信号传导过程[16]。HUA1 属于CCCH 型锌指蛋白,在拟南芥花发育过程中,HUA1 参与了RNA 结合蛋白AGAMOUS 前mRNA 的加工[17]。在矮牵牛中,PetSPL3可通过控制特定细胞类型的细胞分裂和/或扩增,参与各种营养器官和生殖器官的发育调控,推测矮牵牛中的几种花药特异性锌指蛋白均参与花药生殖组织和非生殖组织中配子的调控[17]。
3.1.3 锌指蛋白参与植物叶片衰老
植株叶片衰老是由其生长发育环境和和体内调控网络主导的。孔照胜[16]研究发现一个新的CCCH 型蛋白OsDOS,该蛋白有2 个CCCH 锌指结构域[14],OsDOS-GFP 融合蛋白定位于细胞核,RNAi(RNA interfere)抑制OsDOS 在叶片自然衰老、果实发育过程中的表达,加速叶片衰老进程。OsDOS过表达植株表现严重的发育迟滞和明显的叶片衰老延缓,以此推测OsDOS作为植物叶片衰老过程的负调控因子。在OsDOSRNAi株系中茉莉酸信号通路的基因表达显著上调,而在OsDOS过表达株系中这些基因表达抑制或变化不明显,表明茉莉酸信号通路可能参与了OsDOS 介导的对叶片衰老的调控。此外,OsDOSRNAi 过表达株系的种子和离体叶片对茉莉酸甲酯的处理分别表现出过度敏感和低敏感反应,进一步表明该蛋白很可能通过调控发育信号和茉莉酸信号通路的整合来延缓水稻叶片衰老。通过对棉花细胞壁再生SSH 库中分离出来的CCCH型串联锌指基因GhTZF1 进行鉴定,该基因主要在早期细胞壁再生过程中表达,在各种营养和生殖组织中也有表达。在干旱胁迫条件下,转基因植物中的超氧化物歧化酶和过氧化物酶的活性明显高于野生型植物,在干旱和茉莉酸甲酯(MeJA)处理下,转基因植物中H2O2的量减少,实时定量PCR分析结果表明,GhTZF1的过表达降低了氧化相关衰老基因的表达。通过在H2O2处理期间转基因植物中改变的一组抗氧化剂基因表明,GhTZF1的过表达也增强了氧化应激耐受性,因此,GhTZF1可以通过调节活性氧的稳态来介导植株干旱胁迫耐受性和叶片衰老[18]。
3.1.4 锌指蛋白参与植物细胞凋亡
植物细胞程序化死亡(PCD)是由复杂的信号转导网络控制的,转录因子在调节植物防御基因表达和抗性反应上发挥着关键作用。C2C2型锌指蛋白在拟南芥中占转录因子总数的6.78%,在水稻中占4.67%,由GATA、CO-like、Dof、YABBY 和 类LSD1 (lesions stimulating disease resistance 1)共5 个基因家族组成,除GATA 外其他都是植物所特有的基因家族[19],其中类LSD1 基因家族编码一类特殊的C2C2型锌指蛋白,结构域一般由25个保守的氨基酸残基组成。研究发现拟南芥LSD1 和LOL1(LSD-One-Like 1)均参与植物PCD 的调控,对从拟南芥类LSD1 中克隆出的AtLSD1和AtLOL1进行功能验证发现:AtLSD1基因是作为植物PCD的负调节因子,限制植物对外界刺激的反应以及后期细胞死亡的扩展,而AtLOL1是植物PCD 的正调节因子,在拟南芥中过量表达可降低植株对毒性病菌的敏感性。对从水稻类LSDI 中克隆出的OsLSDI进行初步探究发现:该基因可作为植物愈伤组织分化的正调节因子和植物PCD的负调节因子。目前发现半胱天冬蛋白酶(caspase)在动物PCD 过程中起主导作用,在植物中还未发现其直系同源蛋白,但发现一类metacaspase 蛋白酶,与其结构高度相似,对metacaspase 基因的结构和主要功能进行初步探究,发现29个植物物种均含有metacas‐pase 基因[20]。根据差异表达分析结果推测拟南芥中9 个metacaspase 基因不同程度地参与了细胞对外界刺激反应的过程,且存在于不同的功能模块内,在生物学功能上也存在一定差异。在拟南芥和水稻中均有2 个含有metacaspase p20 亚基的蛋白质,p20 亚基能够独立引起细胞凋亡[21-22],可基本判定这4 个蛋白质都参与了植物PCD 的调节。通过进一步的研究,发现此类相关基因不仅在植物PCD 中发挥作用,同时也参与细胞物理、化学、生物因子等对外刺激的反应过程[23]。此外杆状病毒凋亡重复序列(BIR)包含CCHC配位基序,保守结构域核心由中央三链β片段和4 个短α螺旋组成,许多BIR结构域蛋白都可通过抑制胱天蛋白酶而参与程序性细胞死亡的调控[23]。
3.2 锌指蛋白参与植物胁迫调控
由于外界自然环境条件复杂多变,植物在其生长发育过程中经常遭受非生物因素和生物因素等逆境胁迫,导致植株生长受到抑制、损害、作物产量下降甚至死亡,影响农业生产和生态环境。目前关于锌指蛋白的研究为其参与植物胁迫耐受性机理提供了依据,并为改善植物的胁迫耐受性提供了重要的方法。
3.2.1 参与植物的非生物胁迫调控
通过PCR辅助方法从拟南芥中克隆了锌指蛋白基因家族的4个不同成员,一个与已经报道的基因STZ/ZAT10相同,其余3 个为尚未鉴定的基因,将其命名AZF1(拟南芥锌指蛋白1)、AZF2和AZF3。AZF 和STZ 编码的蛋白质与两个手指的半胱氨酸的其他成员一样,都包含两个典型的半胱氨酸锌指图案,由长的垫片隔开,在锌指基序之外还识别出3 个保守区域,分别称为B-box,L-box 和DNLbox,这4 个基因位于锌指基序序列构建的系统树的同一分支上,进一步表明了它们的结构和功能关系。通过RNA 印迹分析表明:4 个基因主要在根中表达,并在其他器官中也有不同水平表达;4 个基因的表达均对水分胁迫有反应,高盐处理会导致这些基因的表达水平升高,低温处理增加了AZF1、AZF3和STZ表达的水平, ABA 处理可诱导AZF2表达,其中诱导的时间及过程与高盐诱导的时间过程相似(ABA 可控制植物对干旱和盐胁迫的响应,包括气孔关闭、与渗透调节有关的基因激活[13])。原位定位表明AZF2在盐胁迫条件下,mRNA 积累在根的伸长区中。这些结果表明AZF1、AZF2、AZF3和STZ它们都以ABA 依赖性或非依赖性途径参与调控水分胁迫的下游基因(表2)[24]。
OsTZF1是水稻CCCH型锌指家族的一员,干旱、高盐胁迫以及过氧化氢诱导了OsTZF1基因的表达,OsTZF1基因的表达也可被脱落酸、茉莉酸甲酯和水杨酸诱导。在水稻胚芽鞘、幼叶、愈伤组织和穗组织中均观察到含有OsTZF1启动子与β-葡萄糖醛酸酶融合组织的化学活性。有压力存在时细胞质中可观察到OsTZF1绿色荧光蛋白定位。结合RNA测定表明OsTZF1可能与应激反应基因的RNA 代谢有关。OsTZF1的转基因水稻植株表现种子萌发延迟、苗期生长缓慢和叶片衰老延迟症状,与对照相比,RNAi 的植株表现为种子萌发早、幼苗生长快、叶片衰老早。OsTZF1在植物中过度表达(OsTZF1-OX)时,植物对干旱和高盐胁迫有很好的耐受性,反之亦然,OsTZF1可以作为一种生物技术工具,通过控制胁迫响应基因的RNA代谢来改善各种植物的胁迫耐受性[25]。
从水稻中分离克隆得到OsCOIN转录因子,观察发现OsCOIN在转基因水稻品系中的过表达显著增强了植株对寒冷、盐和干旱胁迫的耐受性,细胞脯氨酸水平随着OsP5CS表达的上调而增加[26]。气孔控制CO2的吸收并优化水分利用效率,H2O2是诱导气孔关闭的重要信号分子,通过克隆并鉴定DST(抗干旱和耐盐性)锌指转录因子,发现它通过直接调节H2O2相关基因的表达进而负调控气孔关闭。DST 功能丧失会促使气孔关闭,降低气孔密度,从而提高水稻的抗旱性和耐盐性。
3.2.2 参与植物的生物胁迫调控
由水稻黄单胞菌(Xoo)引起的细菌性枯萎病是全世界范围内最严重的水稻病害之一,先前的研究表明[27],锌指蛋白家族成员水稻cDNA EI38D7(GenBank 登录号BF108310)是防御反应基因,它的表达在水稻对Xoo的抗性中被诱导(表3)。
从接种无毒黄单胞菌辣椒叶中分离出编码C2H2型锌指转录因子的辣椒锌指蛋白基因CaZFP1,该CaZFP1蛋白是核靶向蛋白质,全长没有转录激活活性,其功能是作为转录调节,而CaZFP1的C 端区域具有反激活活性。该CaZFP1转录因子在胡椒茎、根、花和红色果实中均表达,但在叶和绿色果实中未能检测到。CaZFP1响应于炭疽菌,Ca‐ZFP1转录基本定位于辣椒叶中脉中维管束的韧皮部细胞,非生物激发子和环境胁迫可以更早地诱导CaZFP1基因。在转基因拟南芥植物中过表达CaZFP1基因不仅增强了对丁香假单胞菌PV 感染的抵抗力还增强了番茄的耐旱能力[59]。
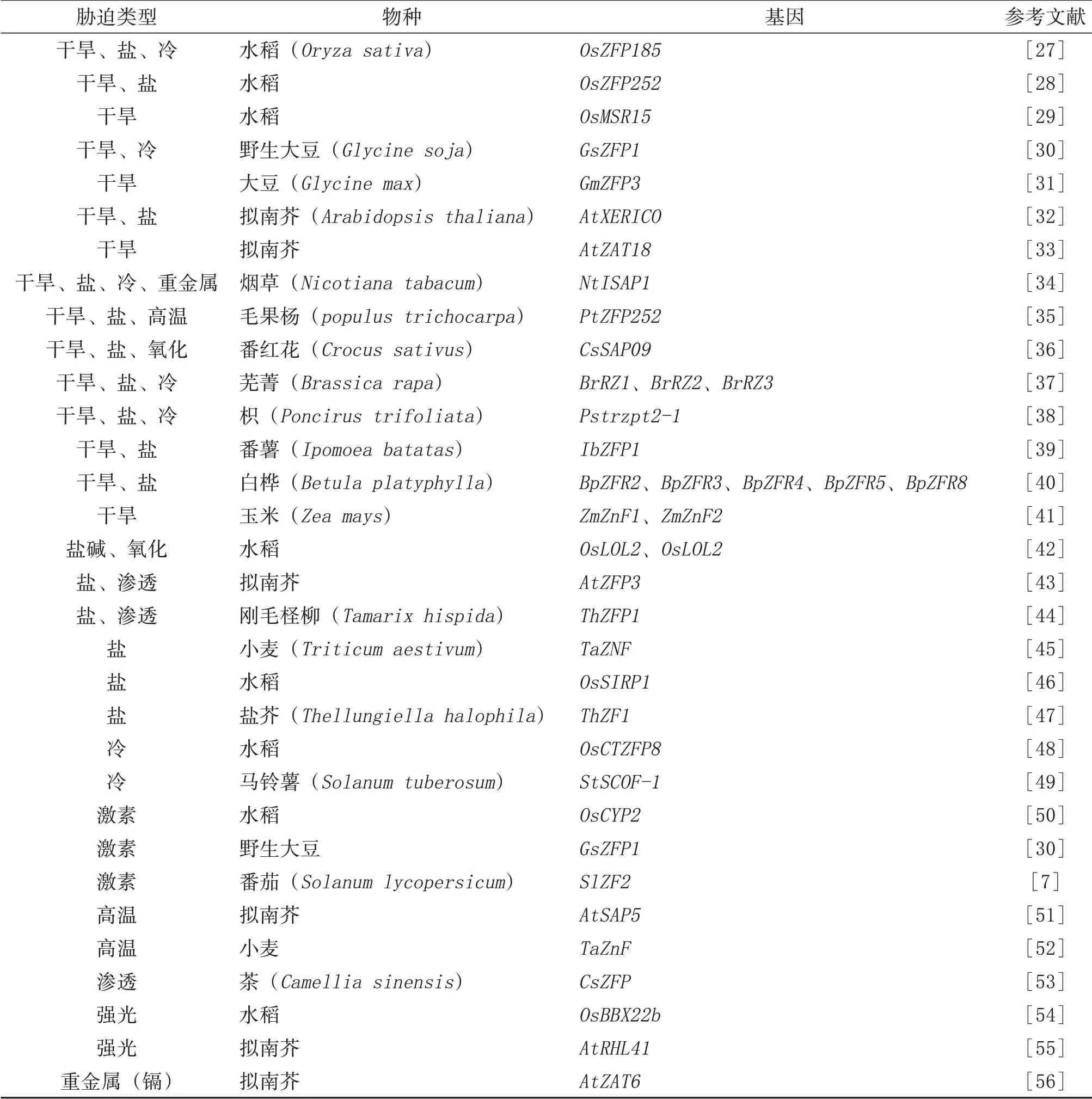
表2 锌指蛋白转录因子在非生物胁迫中的调控作用

表3 锌指蛋白转录因子在生物胁迫中的调控作用
4 锌指蛋白的转录调控机制
非洲爪蟾的5S 基因特异性转录因子TFIIIA 中的九个串联锌指重复序列与5S DNA和5S核糖体RNA特异性结合,进一步说明了锌指蛋白可与DNA 或RNA 进行特异结合。TFIIIA 与DNA 结合的机制较为复杂,氨基末端的3 个锌指识别一个13 bp 的序列并结合在主沟中,这些指状结构主要在DNA的一条链上和鸟嘌呤进行接触。α螺旋表面上的特定组氨酸(非锌配位其残基)和精氨酸残基参与了DNA识别,紧接在α螺旋之前和在螺旋位置2、3 和6 处(在保守的组氨酸之前)的带电荷氨基酸参与DNA鸟嘌呤的氢键结合。TFIIIA 的第2 指在这些DNA 接触位置上含有组氨酸和精氨酸残基,并发现TFIIIA 的第4 指至第7 指贡献了TFIIIA 结合5S rRNA 的95%的自由能,而第1~3 指在结合启动子方面做出了类似的贡献。此外,这些锌指中的每一个都最低限度地识别序列GGG,诱变实验还显示了这些氨基酸在指定DNA识别中的重要性[6]。
锌指蛋白不仅可以识别DNA 和RNA,还能与DNA-RNA 杂交双链分子及其他锌指蛋白或自身结合,在转录和翻译水平上调控基因的表达。DNA 结合锌指蛋白的3 个不同基序为:锌指(i)、锌簇(ii) 和锌扭曲(iii)[61]。酵母转录激活因子(GAL4)和DNA 结合结构域是第1 个被确定的,该蛋白内包含两个锌结合位点,并且在蛋白质中半胱氨酸残基是唯一的锌配体。在GAL4 中,两个锌原子结合6个半胱氨酸残基,形成与金属硫蛋白类似的“锌簇”,GAL4 的两个锌原子之间的距离约为3.5 A,在糖皮质激素受体中,每个锌原子与4 个半胱氨酸残基结合,“锌扭曲”由位于两个锌原子之间的螺旋DNA识别位点表示。在包含多个锌原子的DNA结合蛋白中,锌簇和锌扭曲参与调节基础代谢、次生代谢、减数分裂等过程。
5 展望
锌指在结构上是多样化的,它存在于蛋白质一级结构中,在细胞增殖和代谢、转录和翻译、信号转导和凋亡等过程中发挥广泛的功能。锌指在与多种化合物,如核酸、蛋白质和小分子等结合过程中起着交互模块的作用,其结构研究为其功能的多样性和特异性提供了新的依据。目前已经确定的大量推定的锌指图案中,只有少数在结构上得到了表征,要阐明锌指蛋白的结构、功能和作用机理还需要继续研究。目前已经明确的是,这些小的、独立折叠的蛋白质结构域在调节植物的生物学功能中起着至关重要的作用,尽管大多数已知的锌指蛋白都是转录调节因子,但它们也可以参与染色质重塑,RNA 结合或蛋白-蛋白相互作用这些生物学过程[62]。即使是经典的C2H2锌指基序也不能只简单地看作是核酸结合基序,还要看到其完成除DNA、RNA 识别和包装之外的其他功能[23]。
目前该转录因子研究主要体现在克隆及定位分析、组织表达特异性及胁迫条件下的表达规律、转基因植株的抗逆性检测等[7,63],对锌指蛋白除基本逆境胁迫的信号传导及转录因子间的相互作用机制等方面研究较少,如该转录因子通过哪些途径、以何种方式调控下游基因表达、与其它转录因子相互作用的方式及原理等。随着分子生物学的发展和研究水平的提升,以及对该转录因子的研究更加深入全面,有望在利用锌指蛋白基因资源改良及培育新品种中发挥重要作用。
猜你喜欢
中学生天地(A版)(2023年1期)2023-02-17 00:33:04
昆明医科大学学报(2022年1期)2022-02-28 07:46:10
实用临床医药杂志(2021年13期)2021-01-10 19:52:18
学苑创造·A版(2020年12期)2020-01-07 14:07:23
中国外汇(2019年15期)2019-10-14 01:00:34
生命科学研究(2018年1期)2018-05-29 01:12:47
上海农业学报(2017年3期)2017-04-10 12:39:14
山东农业工程学院学报(2016年6期)2016-12-01 05:38:19
作文教学研究(2016年1期)2016-07-05 12:22:47
中华胰腺病杂志(2015年5期)2015-12-08 12:18:10
