
玉米锌指蛋白基因ZmLSD1 的生物信息学及表达特性分析
2020-05-15 09:37周开明赵白杨张丙林刘卫娟邹华文
辽宁农业科学 2020年2期
周开明,赵白杨,张丙林,刘卫娟,邹华文
(长江大学农学院,湖北 荆州 434025)
非生物胁迫是威胁植物生长发育的主要环境因素之一,例如干旱、高温、高盐等逆境。 转录因子作为具有调节基因表达功能的调控因子,在植物非生物胁迫响应中发挥重要作用。 锌指蛋白是真核生物中较为丰富的一类转录因子,在植物的生长发育及非生物胁迫调节中发挥重要的调控作用[1~2]。 拟南芥锌指蛋白基因STZ、ZAT18 被证实积极参与植株的干旱胁迫响应[3~4]。 张彬等对谷子锌指蛋白基因SiZFP182 的研究发现,该基因的表达受干旱胁迫的诱导[5]。 在大豆中过量表达锌指蛋白基因SCTF-1,转基因株系的耐冷性得到显著提高[6]。 过量表达玉米锌指蛋白基因ZmAN14 的烟草,其苗期的耐盐性及耐旱性都得到明显提升[7]。 此外,锌指蛋白基因还被发现与多种非生物胁迫相关。 宁露云等研究发现,矮牵牛锌指蛋白基因PhTZF1 的表达受低温、干旱、ABA 和高盐等强烈诱导[8]。 Mukhopadhyay 等研究证实水稻锌指蛋白基因OsSAP1 的表达受低温、高盐、重金属、干旱和激素ABA 等条件的诱导,且过量表达OsSAP1 基因的烟草,其耐冷、耐盐及耐干旱水平显著提高[9]。 而水稻OsZAT12基因的过表达,转基因株系的生长发育会受到影响[10]。
类LSD1 转录因子是一类特殊的C2C2 型锌指蛋白,其特征是含有特殊的锌指结构(zf-LSD)[11]。 自从Dietrich 首次将LSD1 基因克隆后,对LSD1 基因的功能研究就不断被报道[11]。Kliebenstein 等的研究表明LSD1 能通过应答水杨酸信号通路清除体内活性氧,间接抑制细胞死亡,暗示LSD1 在植物逆境响应调控的作用[12]。 Liu等发现LSD1 家族中,含有3 个zf-LSD1 结构域的亚家族成员,其功能性要强于含有1 个或者是2个zf-LSD1 结构域的亚家族成员,主要的表现是能更多的参与植物细胞死亡调控、抗病性和非生物胁迫抗性调节[13]。 对苦荞类LSD1 转录因子的表达分析显示,锌指蛋白基因FtLSD1、FtLOL1 对非生物胁迫的应答强烈[14~15],猜测该基因可能参与苦荞抗逆响应机制。 水稻OsLOL2 在拟南芥中的过表达研究表明,该类LSD1 基因具有提高植物抗盐碱性的功能[16]。
长江大学农学院前期克隆了调控玉米转脂蛋白基因ZmLTP3 表达的锌指蛋白类转录因子[17],并命名为ZmLSD1(GenBank: MH119132. 1)。 该基因长660 bp,编码219 个氨基酸,含有3 个zf-LSD1 结构域,进化关系上与高粱LOL3 蛋白和水稻LOL3 蛋白有较高的亲缘关系。 为深入探究锌指蛋白基因ZmLSD1 的功能,本研究首先对ZmLSD1 进行生物信息学分析,同时运用qRTPCR 技术分析该基因在高温、低温、盐、干旱、脱落酸以及水杨酸处理下的表达特性。
1 材料与方法
1. 1 试验材料
玉米B73 自交系,长江大学农学院保存。
1. 2 试验方法
1. 2. 1种植与处理 选取大小一致的玉米种子,1%次氯酸钠消毒5 min,蒸馏水冲洗3 次,置于28 ℃培养箱催芽。 筛选发芽一致的种子,放入1/4 Hoagland 培养液水培,每7 d 更换2 次培养液,培养至三叶期进行处理。 干旱处理为20% PEG 6000 模拟干旱,盐处理为0.2 mol/L NaCl 溶液,对照组均为正常培养,处理48 h 后取样;低、高温处理分别采用4 ℃、42 ℃光照培养箱,对照组均为正常温度培养,处理4 h 后进行取样;激素处理采用0.1 mol/L 的ABA、SA 进行叶面均匀喷施,对照组喷施蒸馏水,处理48 h 后取样。 取样部位为幼苗的根、胚芽鞘、叶,液氮速冻后保存于超低温冰箱中;每个处理设置3 个生物学重复。
1. 2. 2玉米总RAN 的提取及cDNA 的合成 样品材料液氮低温研磨后,参照宝日医生物技术(北京)有限公司TRIzol 试剂产品说明方法进行总RNA 的提取,1%琼指糖凝胶电泳进行质量检测。 残留DNA 的去除以及cDNA 合成采用全式金公司试剂盒TransScript One-Step gDNA Removal and cDNA Synthesis SuperMix,得到的cDNA 使用Thermo NanoDrop One 进行浓度测定后暂存于-20 ℃。
1. 2. 3玉米ZmLSD1 基因表达量分析 采用玉米GAPDH 基因为内参,ZmLSD1 基因的qRT-PCR上下游引物通过Primer 5.0 软件设计,引物委托上海生工进行合成(表1)。 试验所使用的SYBR Green I 荧光标记染料购于天根生化公司的Real-Universal 彩色荧光定量预混试剂,荧光定量PCR仪型号为bio-rad CFX connet。 反应条件:98 ℃ 5 min;98 ℃ 10 s,62 ℃ 30 s;40 个循环。 每个样品设置2 个技术重复,相对定量的结果采用2-△△Ct 法进行计算基因的相对表达量。
1. 2. 4玉米ZmLSD1 基因的生物信息学分析 在线工具Expasy ProtParam(https:/ /web. expasy.org/protparam/)被用于对ZmLSD1 基因蛋白特性进行预测分析,DNAMAN 软件进行序列比对后使用MEGA-X 软件进行进化树的构建,使用在线工具InterProscan[18](http:/ /www. ebi. ac. uk/interpro/search/sequence-search)分析ZmLSD1 蛋白结构域,蛋白二级结构预测分析则采用在线工具 SOPMA (https:/ /npsa-prabi. ibcp. fr/cgi-bin/npsa_automat. pl? page=npsa_sopma. html),TMHMM Server v. 2. 0(http:/ /www. cbs. dtu. dk/services/TMHMM/) 被用于跨膜结构分析,ProtScale(https:/ /web. expasy. org/protscale/) 用于分析ZmLSD1 蛋白的亲疏水性分析,NetPhos 3.1 (http:/ /www. cbs. dtu. dk/services/NetPhos/)则用于进行磷酸化位点预测分析。 Softberry 在线工具protcomp9.0(http:/ /www. softberry. com/berry. phtml? topic=protcomppl&group=programs&subgroup=proloc)进行基因的亚细胞定位。
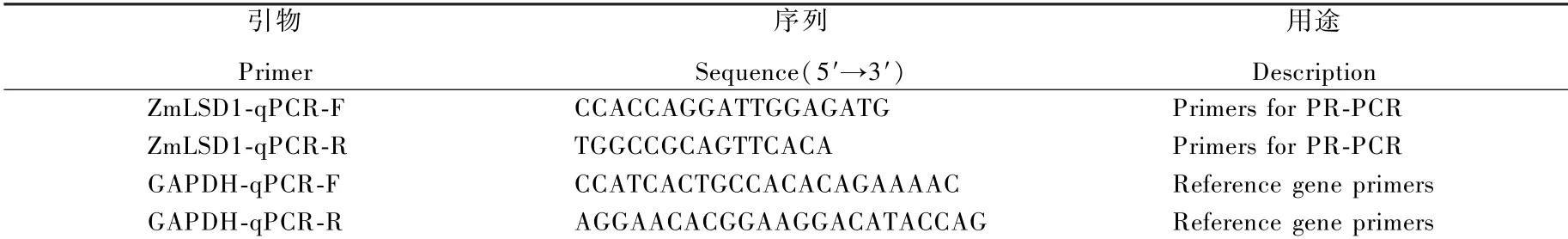
表 1 引物序列及用途Table 1 Primers used in this study
2 结果与分析
2. 1 玉米ZmLSD1 的蛋白序列分析
ProtParam 在线分析显示,ZmLSD1 基因共编码 219 个 氨 基 酸, 其 分 子 式 为C978H1607N295O306S23,分子质量23 130 u,理论等电点为8.95,总平均亲水性0. 057,亲水性系数介于-0.5 到0. 5 之间,属于两性氨基酸(图1,A)。NetPhos 3. 1 在线分析显示,ZmLSD1 中有20 个Ser 磷酸化位点,9 个Thr 磷酸化位点(图1,B)。
利用InterProscan 对ZmLSD1 蛋白的结构域分析发现,该蛋白中含有3 个zf-LSD1 结构域,分别位于氨基酸51-75、90-114 和128-152 位置。SOPMA 预测结果显示,ZmLSD1 的二级结构中含有的α 螺旋约占10.05%,β 转角约占7.31%,延伸链约占30.14%,其余的115 个氨基酸为无规则卷曲(图1,C)。 Softberry 在线工具进行的蛋白亚细胞定位预测结果显示,该蛋白最大可能性位于细胞核。
2. 2 玉米ZmLSD1 的蛋白同源性及进化树分析
在NCBI 蛋白序列比对的结果表明,ZmLSD1编码的氨基酸序列与高粱(Sorghumbicolor)LOL3蛋白、水稻(OryzasativaJaponica Group)LOL3 蛋白和小麦(Triticumaestivum)LOL2 蛋白序列相似度较高,相似性分别达到了94.83 %、77.96 %和73.60 %。 用DNAMAN 软件将ZmLSD1 蛋白序列与其它物种进行蛋白同源性比对分析(图2),结果显示ZmLSD1 与高粱、水稻、小麦、荞麦以及拟南芥的锌指蛋白存在类似结构,其序列中均含有3 个保守的zf-LSD1 结构域,表明ZmLSD1 属于典型的锌指蛋白家族,推测它们具有相似的功能。
基因的进化关系分析显示(图3),ZmLSD1与高粱LOL3、水稻LOL3、水稻LOL2、玉米LOL1和小麦LOL2 的亲缘关系最近。 进化树主要分为3 个分支,LSD1-like 分支中聚集了包括ZmLSD1在内的类LSD1 基因,而拟南芥AtSZF1、水稻Os-DOS 和小麦TaTZF1 共同聚集于CCCH 分支,第三分支(C2H2) 则聚集了棉花GaZFP、拟南芥AtAZF2 和水稻OsZFP182 基因。 进化树成员的氨基酸序列分析发现,不同分支的锌指蛋白含有不同的典型结构域,LSD1-like 分支上LSD1 和LOL 类蛋白含有典型的zf-LSD1 锌指蛋白结构域,而C2H2 分支和CCCH 分支分别含有典型的zf-C2H2 和zf-CCCH 结构域。
2. 3 玉米ZmLSD1 的表达特征分析
2. 3. 1ZmLSD1 的组织表达分析 采用荧光定量PCR 技术分析ZmLSD1 基因在不同部位中的表达情况。 结果表明(图4),ZmLSD1 的在叶中的表达量最高,其次是胚芽鞘,在根中的表达水平相对较低;该基因在叶中的表达量是根中表达量的3. 18 倍,在胚芽鞘中表达量是根中表达量的1.55 倍。 推断ZmLSD1 基因参与调控表达的部位以叶为主。
2. 3. 2ZmLSD1 在不同处理下的表达分析 利用qRT-PCR 分析ZmLSD1 基因在6 种非生物处理下不同植株部位的表达水平。 根部位的结果如图5 显示,ZmLSD1 基因受低温、高温、高盐、干旱和ABA 处理的诱导而上调表达,其中低温和干旱处理后与对照的相对表达量分别为5. 47、18. 4,达到差异极显著水平;水杨酸处理下该基因的表达量则降低。 胚芽鞘的检测结果表明(图6),ZmLSD1 基因在盐、干旱以及2 种激素(ABA 和SA)的处理下呈现上调表达趋势,其中干旱胁迫下的相对表达量为8. 21,差异水平极显著;而4 ℃和42 ℃温度处理下该基因的表达受抑制。ZmLSD1 在叶中的表达受盐、干旱和ABA 的影响,其中干旱处理产生的影响最大,而在低温、高温和水杨酸条件下该基因表现为下调表达(图7)。 实验结果显示,ZmLSD1 基因对干旱、盐和ABA 胁迫存在正向响应,而低、高温以及水杨酸环境能抑制该基因的表达,但在不同部位中的敏感程度还是存在一定的差异;该基因在根中对干旱、高温和低温最敏感,在胚芽鞘中的表达受干旱和低温的影响程度最高,在叶中则对盐和干旱敏感。 结果表明ZmLSD1 的表达受干旱、盐、高温和低温胁迫的诱导,推断该基因可能参与干旱、盐、高温和低温等非生物胁迫的响应。
3 结论与讨论
干旱、盐等非生物胁迫严重影响作物的生长发育,是限制作物产量的主要因素之一[19]。 研究发现锌指蛋白转录因子在植物的生长发育以及逆境胁 迫 下 的 调 控 表 达 发 挥 着 重 要 作 用[4,20~21],LSD 类锌指蛋白广泛参与植物生物及非生物逆境信号传导过程[22],其典型的特征是形成锌指结构并在蛋白质调控中发挥作用[23]。 大量研究表明,含有3 个zf-LSD1 结构域的锌指蛋白基因在逆境响应过程中发挥重要作用[14~16,24];ZmLSD1 也含有3 个zf-LSD1 结构域,进化分析发现其与水稻OsLSD1、玉米ZmLOL1、拟南芥AtLSD1 等蛋白有较高的同源性,属于典型的锌指蛋白家族成员。
本研究发现玉米锌指蛋白基因ZmLSD1 受植物非生物胁迫的诱导表达,如干旱、盐、高温和低温等。 在干旱和盐处理后,ZmLSD1 基因在根、胚芽鞘和叶中的表达量均呈现上调表达趋势,说明该基因的表达受干旱和盐的诱导。 这与香蕉MaZAT10 基因的表达模式一致,香蕉C2H2 型锌指蛋白基因MaZAT10 受干旱、盐和低温处理的诱导而显著上调表达[25]。 基于温度(4 ℃、42 ℃)处理的研究发现,ZmLSD1 基因在胚芽鞘和叶部位的转录表达水平受抑制。 该基因对冷、热胁迫的响应与苦荞FtLOL1、水稻OsBBX6 存在一定的相似性,苦荞锌指蛋白基因FtLOL1 在4 ℃的低温胁迫和水杨酸处理下表达量降低[15],水稻锌指蛋白基因OsBBX6 则被发现负向响应高温胁迫[26]。ZmLSD1 基因在脱落酸处理下上调表达,说明ZmLSD1 基因可能是通过ABA 信号路径介导植物的逆境响应。 而在水杨酸的处理下,ZmLSD1基因在根和叶部位中的表达量下降。 这与拟南芥锌指蛋白基因AtLSD1 的表达趋势相反,AtLSD1基因通过积极响应SA 信号,提升活性氧的清除以降低非生物胁迫的伤害[27]。
玉米锌指蛋白基因ZmLSD1 对不同的非生物处理表现出不同的表达特征,可能是该转录因子的DNA 结合差异或蛋白互作差异产生的结果,说明ZmLSD1 与玉米逆境响应的密切相关,但其确切的生物学功能,有待于进一步研究。
猜你喜欢
湖北农业科学(2022年11期)2022-07-18
军事文摘(2021年16期)2021-11-05
今日农业(2020年23期)2020-12-15
实用肿瘤学杂志(2020年4期)2020-12-08
劳动保护(2018年8期)2018-09-12
山东工业技术(2016年15期)2016-12-01
飞碟探索(2015年9期)2015-11-05
红领巾·探索(2015年9期)2015-09-10
医学综述(2011年12期)2011-12-09
亚热带农业研究(2011年3期)2011-09-29
