
3种紫堇属植物叶片光合特性研究
2020-04-16 03:35陈晓英郭晓云秦双双张占江
植物资源与环境学报 2020年1期
陈晓英, 李 翠, 郭晓云, 秦双双, 张占江
(广西壮族自治区药用植物园 广西药用资源保护与遗传改良重点实验室, 广西 南宁 530023)
岩黄连(CorydalissaxicolaBunting)隶属于罂粟科(Papapveraceae)紫堇属(CorydalisDC.),中文学名石生黄堇,是治疗肝脏类疾病的常用壮药和瑶药[1-2],多生长在石山地区峭壁的石缝中,采挖困难,目前其野生资源处于濒危状态[3-4],因此,一方面需要对岩黄连进行引种驯化和人工栽培[5-7],另一方面需要挖掘岩黄连同属替代植物资源。小花黄堇〔C.racemosa(Thunb.) Pers.〕和北越紫堇(C.balansaePrain)也隶属于紫堇属,其生物碱成分与岩黄连相似[8-11],有望替代岩黄连入药,从而缓解岩黄连药材供应不足的现状。但是,关于小花黄堇和北越紫堇的生长和生理特性及生态适应性却知之甚少,不利于大规模栽培生产。
研究者根据植物光合特性不但能够快速了解植物的生长和生理特性,而且能够根据各光合作用参数的变化分析植物对光照、水分、温度和CO2浓度等的需求,明确植物对生态环境的适应性[12-15]。目前,关于岩黄连、小花黄堇和北越紫堇光合特性方面的研究报道甚少,仅见韦记青等[16]对岩黄连的光合与蒸腾特性及其对光照强度和CO2浓度的响应进行了研究,因此,为了这3种植物的大规模人工种植,应尽快开展其光合特性方面的全面研究。
鉴于此,笔者对岩黄连、小花黄堇和北越紫堇叶片的光合色素含量、光合特征参数光响应曲线及净光合速率光响应曲线特征参数进行了比较,并对这3种植物的光合型进行了分析,以探明这3种植物的光能利用特性和生态适应性,并为岩黄连及其同属植物的引种驯化和生态栽培提供理论基础和科学依据。
1 材料和方法
1.1 试验地概况
试验地位于广西壮族自治区药用植物园温室大棚内(东经108°19′、北纬22°51′)。该区域属亚热带季风气候,年均温17 ℃~23 ℃,年降水量1 250~2 800 mm,年均日照时数1 827 h,无霜期334 d。
1.2 材料
以半野生栽培(对野生的成苗植株进行常规水肥等人工栽培管理措施)的小花黄堇、北越紫堇和岩黄连为实验材料,经广西壮族自治区药用植物园标本馆黄宝优高级工程师鉴定。3种植物分别种植在温室大棚内的不同区域,于2017年9月至11月(营养生长期)分别选取植株最顶端叶片进行光合色素含量测定,并绘制光合特征参数光响应曲线。
1.3 方法
1.3.1 光合色素含量测定 参照黎修东等[17]的方法测定叶片的光合色素含量。每种植物选取5株植株,将采集的叶片剪碎后混匀;称取0.2 g碎叶,加入体积分数95%乙醇,置于黑暗处直接浸提,待叶片变白后定容至50 mL,即为光合色素提取液;用752N紫外分光光度计(上海仪电分析仪器有限公司)分别在波长665、649和470 nm处测定该提取液的吸光值,并计算叶绿素a(Chla)、叶绿素b(Chlb)、类胡萝卜素(Car)和总叶绿素(Chla+Chlb)的含量以及叶绿素a含量与叶绿素b含量的比值(Chla/Chlb)。每个指标重复测定3次。
1.3.2 光合特征参数光响应曲线绘制 在晴天8:30至11:30,每种植物选取3~5株植株,使用LI-6400XT便携式光合作用测量系统(美国LI-COR公司)测定植株最顶端叶片的净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)和胞内CO2浓度(Ci),并计算瞬时羧化效率(Pn/Ci)[18]、气孔限制值(Ls)[19]和水分利用效率(WUE)[20]。测定时,首先进行充分的光诱导,然后设置CO2浓度400 μmol·mol-1、空气相对湿度60%、叶温35 ℃。用LI-6400XT红蓝光源(美国LI-COR公司)控制光合有效辐射(PAR),设定PAR梯度依次为1 500、1 200、1 000、800、600、400、200、150、100、50、20和0 μmol·m-2·s-1,每个PAR梯度的适应时间为3~5 min。根据测定结果绘制各光合特征参数的光响应曲线。
1.3.3 净光合速率光响应曲线特征参数分析 采用改良的直角双曲线模型[21-22]拟合3种紫堇属植物Pn值的光响应曲线,据此获得Pn值光响应曲线的特征参数,包括表观量子效率(AQY)、最大净光合速率(Pmax)、光补偿点(LCP)、光饱和点(LSP)和暗呼吸速率(Rd)。
1.3.4 光合型分析 根据何亚婷等[23]和唐海萍等[24]的方法,利用与光合作用过程直接相关的Pn值、Tr值、气温与叶温的差值和Gs 值计算光合型判别标准值(Gx),根据Gx值初步判断3种紫堇属植物的光合型。Gx≥0表示植物光合型为C3型,Gx<0表示植物光合型为C4型。
1.4 数据处理及统计分析
采用SPSS 24.0统计分析软件计算数据的平均值和标准误,对各光合特征参数的光响应曲线进行非线性拟合并制图。
2 结果和分析
2.1 叶片光合色素含量比较
根据3种紫堇属植物叶片光合色素含量的比较结果(表1),岩黄连叶片的叶绿素a、叶绿素b和总叶绿素含量均最高,北越紫堇次之,小花黄堇最低;小花黄堇叶片的叶绿素a含量与叶绿素b含量的比值最高,岩黄连次之,北越紫堇最低;岩黄连叶片的类胡萝卜素含量最高,小花黄堇次之,北越紫堇最低。与岩黄连相比,小花黄堇和北越紫堇叶片的叶绿素a含量分别减少了14.21%和13.71%,叶绿素b含量分别减少了28.85%和4.81%,总叶绿素含量分别减少了19.27%和10.30%,叶绿素a含量与叶绿素b含量的比值分别增加了21.16%和减少了7.94%,类胡萝卜素含量分别减少了9.30%和18.60%。


1)Chla: 叶绿素a含量 Chlorophyllacontent; Chlb: 叶绿素b含量 Chlorophyllbcontent; Chla+Chlb: 总叶绿素含量 Total chlorophyll content; Chla/Chlb: 叶绿素a含量与叶绿素b含量的比值 Ratio of chlorophyllacontent to chlorophyllbcontent; Car: 类胡萝卜素含量 Carotenoid content.
2.2 叶片光合特征参数光响应曲线比较
根据3种紫堇属植物光合特征参数的光响应曲线(图1),3种紫堇属植物叶片净光合速率的光响应曲线均随着光合有效辐射(PAR)增大先持续升高后趋于平稳。在弱光(PAR<400 μmol·m-2·s-1)条件下,小花黄堇和北越紫堇叶片的净光合速率与岩黄连的差距较小,但在400 μmol·m-2·s-1≤PAR≤1 500 μmol·m-2·s-1光照条件下,随着PAR值增大,二者叶片的净光合速率与岩黄连的差距逐渐增大,二者叶片的净光合速率均值分别较岩黄连增加了42.96%和30.97%。3种紫堇属植物叶片瞬时羧化效率的光响应曲线与净光合速率光响应曲线相似,但在400 μmol·m-2·s-1≤PAR≤1 500 μmol·m-2·s-1光照条件下,小花黄堇和北越紫堇叶片的瞬时羧化效率均值分别较岩黄连增加了48.63%和34.67%。
由图1可见:岩黄连叶片气孔导度和蒸腾速率的光响应曲线与小花黄堇和北越紫堇明显不同。随着PAR值增大,岩黄连叶片的气孔导度和蒸腾速率均缓慢升高,而小花黄堇和北越紫堇叶片的气孔导度和蒸腾速率则先缓慢升高后大幅升高。在弱光(PAR<400 μmol·m-2·s-1)条件下,小花黄堇和北越紫堇的气孔导度和蒸腾速率与岩黄连的差距较小,但在400 μmol·m-2·s-1≤PAR≤1 500 μmol·m-2·s-1光照条件下,随着PAR值增大,二者叶片的气孔导度和蒸腾速率与岩黄连的差距逐渐增大,二者叶片的气孔导度均值分别较岩黄连增加了33.17%和53.08%,二者叶片的蒸腾速率均值分别较岩黄连增加了33.39%和10.98%。


图1 3种紫堇属植物叶片光合特征参数的光响应曲线
Fig. 1 Light response curve of photosynthetic characteristic parameters of leaves of three species inCorydalisDC.
由图1还可见:3种紫堇属植物叶片胞间CO2浓度、气孔限制值和水分利用效率的光响应曲线相似。随着PAR值增大,三者叶片的胞间CO2浓度均先持续下降后趋于平稳,且在三者间的差距一直较小;而三者叶片的气孔限制值和水分利用效率则均表现为先持续升高后趋于平稳,且在400 μmol·m-2·s-1≤PAR≤1 500 μmol·m-2·s-1光照条件下,随着PAR值增大,小花黄堇和北越紫堇叶片的气孔限制值和水分利用效率与岩黄连的差距越来越小。在400 μmol·m-2·s-1≤PAR≤1 500 μmol·m-2·s-1光照条件下,小花黄堇和北越紫堇叶片的胞间CO2浓度均值分别较岩黄连减少了6.09%和3.81%,二者叶片的气孔限制值均值分别较岩黄连增加了21.35%和18.09%,二者叶片的水分利用效率均值分别较岩黄连增加了10.76%和26.47%。
根据岩黄莲、小花黄堇和北越紫堇光合型判别标准值的光响应曲线(图1),这3种紫堇属植物光合型判别标准值均大于10,据此判断这3种植物的光合型均为C3型。
2.3 净光合速率光响应曲线特征参数比较
计算3种紫堇属植物叶片净光合速率光响应曲线的特征参数,结果见表2。由表2可见:小花黄堇的表观量子效率与岩黄连接近,光补偿点较岩黄连减少了21.10%;北越紫堇的表观量子效率较岩黄连增加了25.45%,光补偿点较岩黄连减少了12.57%。小花黄堇的最大净光合速率和光饱和点分别较岩黄连增加了36.34%和31.23%,而北越紫堇的最大净光合速率和光饱和点则分别较岩黄连增加了37.89%和10.59%。此外,与岩黄连相比,小花黄堇和北越紫堇的暗呼吸速率分别增加了71.43%和133.33%。

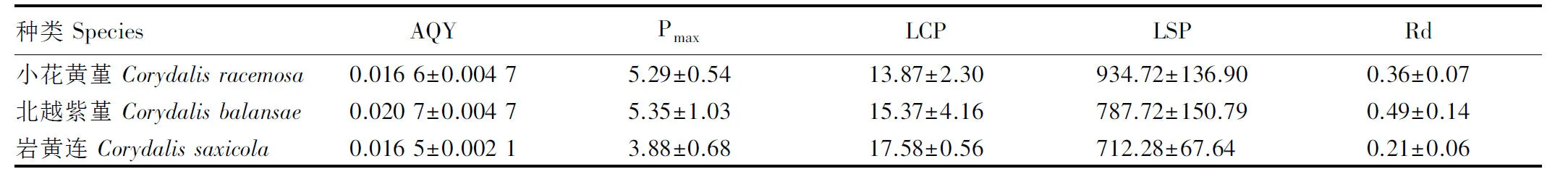
1)AQY: 表观量子效率 Apparent quantum yield (μmol·mol-1); Pmax: 最大净光合速率 Maximum net photosynthetic rate (μmol·m-2·s-1); LCP: 光补偿点 Light compensation point (μmol·m-2·s-1); LSP: 光饱和点 Light saturation point (μmol·m-2·s-1); Rd: 暗呼吸速率 Dark respiration rate (μmol·m-2·s-1).
3 讨论和结论
3.1 3种紫堇属植物光合特性的比较
光合色素主要用于植物对光能的吸收、传递和转化,并执行光保护等功能[25]。虽然光合色素含量与植物的光合特性有关,但变化趋势并非始终一致。小花黄堇和北越紫堇叶片的总叶绿素含量和类胡萝卜素含量均低于岩黄连;在400 μmol·m-2·s-1≤PAR≤1 500 μmol·m-2·s-1光照条件下,随着PAR值增大,小花黄堇和北越紫堇的净光合速率和瞬时羧化效率始终高于岩黄连,且与岩黄连的差距越来越大,说明小花黄堇和北越紫堇的光合碳同化能力高于岩黄连。
本研究结果表明:总体来看,随着光照不断增强,小花黄堇和北越紫堇叶片的气孔导度和蒸腾速率均不同程度升高,且在400 μmol·m-2·s-1≤PAR≤1 500 μmol·m-2·s-1光照条件下,随着PAR值增大,其与岩黄连的差距越来越大;3种紫堇属植物叶片的胞间CO2浓度先持续下降后趋于平稳,而叶片的气孔限制值却先持续升高后趋于平稳,且小花黄堇和北越紫堇叶片的气孔限制值均高于岩黄连。叶片气孔导度和蒸腾速率增加表明小花黄堇和北越紫堇的叶片通过气孔交换了更多的光合作用原料(如CO2和H2O)[26];同时,小花黄堇和北越紫堇叶片气孔导度的增加和胞间CO2浓度的降低导致气孔限制值增加,表明小花黄堇和北越紫堇光合作用的限制因子主要是气孔限制因子,而非电子传递受阻、叶肉细胞羧化阻力、1,5-二磷酸核酮糖(RuBP)再生限制等非气孔限制因子[12,27-28]。此外,虽然小花黄堇和北越紫堇叶片的蒸腾速率增加但同时伴有水分利用效率提高,说明小花黄堇和北越紫堇能将叶片散失的水分更有效地用于光合作用[29-32]。综合上述研究结果,小花黄堇和北越紫堇的光合作用效率较岩黄连高,在相同环境条件下能积累更多的碳同化产物。
相关研究结果[33]表明:植物的表观量子效率和光补偿点能够反映植物对弱光的利用能力,而最大净光合速率和光饱和点则能够反映植物对强光的利用能力。表观量子效率越高,光补偿点越低,表明植物对弱光的利用能力越强;最大净光合速率和光饱和点越高,表明植物对强光的利用能力越强。根据3种紫堇属植物叶片净光合速率光响应曲线特征参数的比较结果,小花黄堇和北越紫堇的表观量子效率、最大净光合速率和光饱和点均高于岩黄连,光补偿点却低于岩黄连,说明小花黄堇和北越紫堇对弱光和强光的利用能力均高于岩黄连。
总体来看,岩黄连、小花黄堇和北越紫堇光合性能的相似性较高,且小花黄堇和北越紫堇的光合碳同化能力优于岩黄连。然而,这3种植物的最大净光合速率为3.88~5.35 μmol·m-2·s-1,光饱和点为712.28~934.72 μmol·m-2·s-1,说明三者利用强光的能力和光合潜力均不高。
3.2 适宜3种紫堇属植物生长的环境条件分析
根据植物叶片净光合速率光响应曲线的特征参数可推断植物生长所需的光照条件[34-35]。根据本研究结果,小花黄堇、北越紫堇和岩黄连的光补偿点为13.87~17.58 μmol·m-2·s-1,光饱和点为712.28~934.72 μmol·m-2·s-1,暗呼吸速率为0.21~0.49 μmol·m-2·s-1,均符合阴生植物的光照生态幅范围[36],据此判断这3种植物均为阴生植物。从岩黄连野生资源分布情况来看,其原生境多为林缘、石缝等日照时间不长、光照不强、凉爽的山地环境[37]。结合阴生植物特征,建议在对小花黄堇和北越紫堇进行迁地保护和引种栽培时,采用大棚和盖草等遮阳方式或林下栽培和间作套种等立体种植模式来调控光照强度[38]。
植物叶片的蒸腾速率和水分利用效率变化与植株水分代谢有关,可在一定程度上反映植物光合生产过程与植株耗水特性的关系[39]。蒸腾速率可反映光合作用对水分的需求以及通过水分运输的矿质养分的供应情况[40]。水分利用效率受微环境影响较大[30,41],可通过基因工程手段予以提高[42-44]。总体来看,岩黄连、小花黄堇和北越紫堇叶片的蒸腾速率和水分利用效率的光响应曲线变化趋势相似,说明这3种植物可能具有相似的水分利用策略。根据岩黄连多生长在南方温暖湿润地区且忌水涝和排水不良[2,11]的特点,在种植小花黄堇和北越紫堇时可以岩黄连自然生长环境中的水分条件为参考控制栽培地水分条件。
3.3 3种紫堇属植物光合型的初步判定
众所周知,高等植物具有C3、C4和CAM 3种光合途径[45]。本研究中,岩黄连、小花黄堇和北越紫堇的光合型判别标准值均高于10,据此判定这3种植物的光合型均属于C3型。这一结论还需要通过观察叶片的维管束鞘细胞及其外侧紧密毗连的1圈叶肉细胞是否共同组成花环解剖结构以及检测其他生理生化指标进行验证。C3型植物缺乏可以提高光合细胞内CO2浓度的Kranz结构,对CO2的同化能力远低于C4型植物[46],但C3型植物体内的碳同化限速酶Rubisco可同时催化底物RuBP与CO2结合的碳还原反应及RuBP与O2结合的碳氧化反应,因此,在岩黄连、小花黄堇和北越紫堇的栽培过程中,可通过增施CO2肥等措施,提高植株叶片的光合效率,从而促进植株体内光合产物的积累。
3.4 结论
综上所述,岩黄连、小花黄堇和北越紫堇的光合特性相似,均属于阴生植物,光合型均为C3型。与岩黄连相比,虽然小花黄堇和北越紫堇叶片的光合色素含量低,但二者对弱光和强光的利用能力更强,光合碳同化能力也更强。
猜你喜欢
阅读(科学探秘)(2020年8期)2020-11-06
农业与技术(2020年12期)2020-07-04
作文新天地(小学版)(2020年5期)2020-06-08
中成药(2019年12期)2020-01-04
绿色科技(2019年2期)2019-05-21
农学学报(2019年1期)2019-02-21
中国果业信息(2019年1期)2019-01-05
新课程·下旬(2017年7期)2017-08-14
小学生导刊(中年级)(2016年9期)2016-05-14
农产品市场周刊(2014年40期)2014-12-19
