
凤翔雍山血池遗址北斗坊7 号坑马骨的古DNA 研究*
2020-04-14 03:38文少卿俞雪儿陈爱东杨苗苗
考古与文物 2020年6期
文少卿 俞雪儿 孙 畅 陈爱东 杨苗苗 袁 靖
(1.复旦大学科技考古研究院;2.复旦大学现代人类学教育部重点实验室;3.陕西省考古研究院)
关于秦汉时期雍地举行祭天的礼仪,在文献中均有记载,但是关于祭祀用牲却很简略。这些马匹是集中豢养专门用于祭祀,还是从全国各地按照一定的标准挑选后用于祭祀呢?通过古 DNA 高通量测序检测,我们将探讨殉马坑内马的来源;同时,利用基因型和表型间的关联性,推断殉马的一些体质特征,如马的性别、毛色、运动能力、步态等。该项研究将为探讨秦汉礼制及历史等提供参考。
一、材料与方法
1.样本采集和提取
本次测试样本源自血池遗址南部的北斗坊地点的7 号长条坑(K7),共有26 个马骨样本。DNA 提取工作在复旦大学现代人类学专业古DNA 实验室进行,遵循标准流程[1]。为最大程度地防止了实验操作带来的污染,我们采用了最新的短片段提取方案[2]。
2.文库构建与测序
为了更加符合高度降解DNA 的特性,我们设计了更为敏感的短扩增子引物系统:其中线粒体扩增子为113~126 bp,SNP 均小于105 bp。具体包括2 个高通量检测Panel(表一):Panel 1(母系线粒体DNA 和性别鉴定),基于该Panel,我们同时测试马DNA 的控制区(15469-16083; 615 bp)和细胞色素b(14215-14549;335 bp),实现了样本母系遗传谱系的确定,以及2 个性别基因(SRY 和AMEL)的测试,鉴定性别。Panel 2(马的生理和体质特征),主要检测5 个方面的相关基因,即毛色、运动能力、步态、身体大小/马肩隆高度和高原适应。
多重PCR 扩增靶向捕获实验的流程包括:靶区域扩增、PCR 产物纯化、第二轮PCR和PCR 产物回收。复合扩增以后,我们通过Agilent 2100 Bioanalyzer 对DNA 文库进行质检,观察接头自连、外援DNA 污染等情况,并对PCR 产物混样测序。
3.数据分析
采用Illumina X10 平台进行双端测序(paired-end sequencing),获取读长为150bp 的测序数据(fastq)。首先通过cutadapt 软件去除接头序列,然后用bwa(version 1.7,参数:mem)将reads 与马参考基因组(Equus caballus 2.0,线粒体参考基因组替换为X79547 序列)进行比对,生成BAM(Binary Alignment/Map)文件。利用samtools 及bcftools 对常染色体目标区域进行基因位点分型检测。对于线粒体和性染色体,我们采用bedtools 从上一步的BAM 文件提取出成功比对到马全基因组的所有reads,再将这些reads 采用bowtie2 软件参考X79547 序列和X、Y 染色体参考序列重新比对,生成新的BAM。对于第二步的BAM 文件,我们应用BAMClipper软件,根据引物物理位置从BAM 文件去除引物序列得到最终BAM 文件。最 后,采 用samtools(version 1.8) 和bcftools(version 1.8)分别对线粒体、X和Y 染色体目标区域进行变异位点的识别,生成VCF(Variant Call Format)文件。

表一 高通量测序Panel 的基本信息
二、结果
1.线粒体DNA 世系鉴定
7 号坑的26 个样本全部获取了DNA 控制区(15469-16083; 615 bp)序列。以 X79547 作为参考序列进行比对。共检测出50 个多态性位点,48 个位点为转换,2 个位点(15670C、16063C)为插入,没有颠换与缺失。此外,这26 个样本中检测出了26 个单倍型,显示出极高的遗传多态性。
根据Jansen 等[3]的关于单倍型类群的划分,26 个单倍型可以归属于6个不同的谱系A~F,进一步可划分到11 个不同的单倍群,分别为A1(4 匹)、A3(1 匹)、A4(1 匹)、A6(1 匹)、B1(2匹)、B2(2 匹)、C2(3 匹)、D2(2 匹)、E(2匹)、F1(3 匹)、F2(5 匹)。
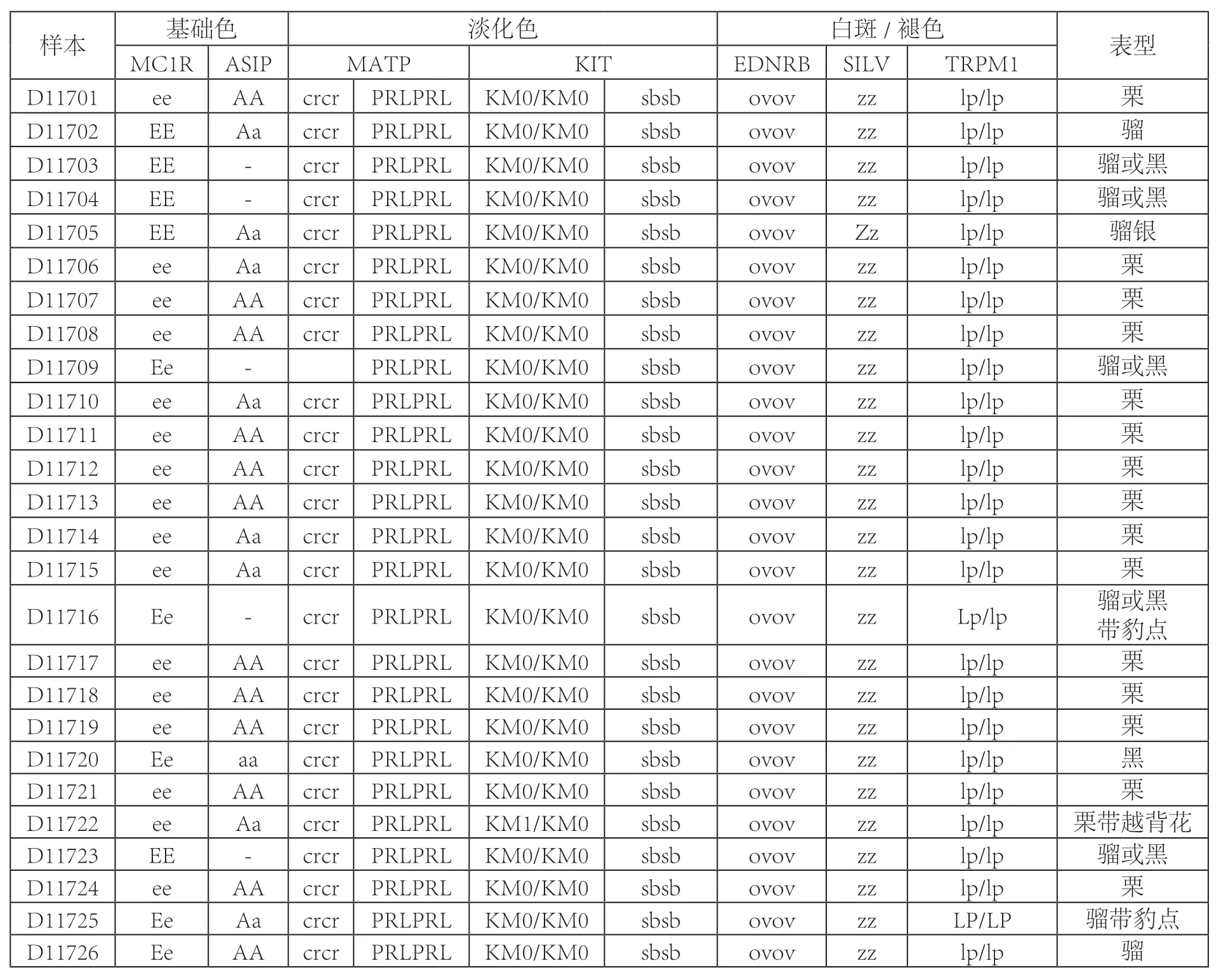
表二 马的毛色基因型与表型特征
2.性别鉴定
根据mapping 在AMEL 和SRY 上reads 的情况,七号坑中公马共计16 匹,母马10 匹,性别比为8:5。
3.体质特征相关位点检测
(1)毛色
马的毛色分为基础毛色、淡化毛色以及白斑/褪色三大类型。基础毛色在马群中比较普遍,可分为骝色、褐骝色、栗色和黑色。一些修饰基因独立或共同发挥作用形成了马的淡化毛色,包括奶酪色、珍珠色、香槟色、银色和暗褐色,以及调控形成更为特殊的毛色表型,如白斑、褪色等,包括分背花色、越背花色、沙边毛色、豹点色等[4]。26 个样本的分型结果为15 个栗色、2 个骝色、1 个黑色、4 个骝色或黑色、1 个骝银色、1 个骝色或黑色带豹点、1个栗色带越背花和1 个骝色带豹点(表二)。
(2)运动能力
对于赛马的全基因组扫描发现,RALGAPA2 基因(chr22:4632335)上携带等位基因G 的马有着更强的运动能力[5],MSTN 基因上位点(chr18:66493737)与最佳赛距相关,携带C/C 基因型的马适合快速、短距离的比赛,C/T 基因型的马在中距离比赛中表现良好,T/T基因型的马耐力更强,更适合长距离奔跑[6]。综合上述位点的分型结果,七号坑26 个样本中除了D11721 和D11723 擅长中距离跑外,其余样本均为T/T 基因型,擅长长距离跑,耐力持久(表三)。
(3)步法
马的行走方式受到了多次人工选择,特别是其交替步法。SNP 位点(chr23:22967656)与马的步态差异有关,DMRT3 基因上的一个突变(chr23:22999655)会显著影响马在完成交替步态时的能力[7]。随后,该突变在全世界的不同马种中进行了验证[8]。在芬恩马(Finn horses)中,携带纯合突变型A/A 的马有着更好的比赛成绩,而携带C 等位基因的马似乎更适合古典骑术比赛[9]。综合位点的分型结果,K7 的26 个样本均携带DMRT3 基因野生型,不太擅长交替步法。
(4)身体大小/马肩隆高度
身高是马最容易观察到的一个表型之一。全基因组关联分析结果表明,位点LCORL(chr3:105547002)、HMGA2 (chr6:81481064)、Z F A T(c h r 9:7 5 5 5 0 0 5 9) 和 L A S P 1(chr11:23259732)联用可以解释高达85%马的体型差异[10]。其中,LCORL (chr3:105547002)位点可以解释18% 的变异[11]。此外,突变(HMGA2:c.83G>A)在矮马中常见,而在正常大小的马的品种中没有发现[12]。综合上述位点的分型结果,七号坑26 个样本非小矮马,除了D11701、D11723 和D11725 身高中等外,其余都偏小,为小型马。
(5)高原适应
生活在高海拔地区会承受生理和代谢方面的挑战,例如缺氧、低温、高辐射和缺乏食物等。Sher L Hendrickson 通过对比低地马和安第斯山马的基因组,发现EPAS1 内含子上的一个SNP位点(rs69 041973;chr15:52570648)与马的高海拔适应相关[13]。七号坑26 个样本中D11706、D11720 和D11725 三个样本为突变型CC,他们可能更能适应高原低氧环境。

表三 马的表型特征相关SNP 分型结果及其表型推测
三、讨论
本次研究的材料为幼马,无法观察一些体质测量和观察类性状,我们试图通过与现代马和古代马的数据相比较,尝试回答血池遗址中祭马来源和是否有严格的挑选标准等问题。
1.祭马来源
为了便于整合已发表文献的数据,本文沿用了Jansen 等[14]在线粒体DNA 单倍群的命名原则。本次检测的26 个样本的母系单倍型完全不同,分属6 个大单倍群(A、B、C、D、E、F)和11 个亚单倍群。
如表四所示,古代家马中[15-21],最常见的世系A(36%),其次依次为世系F(22.5%)、世系E(14.6%)、世系B(9.0%)、世系C(7.9%)和D(6.7%)、世系G(3.4%)。北斗坊七号坑中情况类似,最高频的也是世系A(26.9%)和世系F(30.8%)。如果仅考虑控制区(15494-15740),七号坑的单倍群检出率最高(6/7),单倍型多样性为0.9938±0.0126,说明该遗址的母系来源多样,可能是从各地征集而来。共享单倍型分析同样显示了北斗坊七号坑中葬马在个体水平上的来源多样性。7 号坑中马的多个单倍型分别与位于不同地域的遗址中出土的马的单倍型完全一致(表五):样本D11701、D11702、D11704 显示了与西北地区本地马之间的密切关系;样本D11707、D11711、D11722、D11719、D11725 显示了与蒙古草原马之间的密切关系;样本D11724 显示了与西域马之间的密切关系。
考虑到样本量过少带来的统计学偏差,我们去掉了样本数少于3 个的遗址,并根据线粒体单倍群频率,做了主成分分析。如图一所示,现代马按照地理分组很好地聚集在了一起。关山马、韩国济州马、蒙古马、图瓦马和藏马形成了东亚马的聚类,血池北斗坊7 号坑的古马样本也位于这个聚类之中,并与韩国济州马、蒙古马更近一些。蔡大伟等发现秦公一号大墓出土的古马疑似与阿哈尔捷金马(汗血马)有关[22],在主成分分析图上也得以体现,秦公一号大墓车马坑和板城墓地的古马样本位于阿拉伯马、奥尔洛夫马和阿哈尔捷金马组成的中亚组聚类中。其他古代遗址的样本量均小于5,有较强的统计学偏差,主成分分析的结果不能完全真实地反映其遗传来源。

表四 已检测古代家马的线粒体世系频率分布

表五 北斗坊七号坑样本在中国古代家马中的共享序列分布
2.马的表型特征
(1)性别和毛色
祭祀用马的性别对于理解马在仪式中的含义、马与人的关系、以及古代社会中马的地位和用途至关重要[23]。在已经发表的文献中,祭祀用马一般会有意识地选择使用公马,例如,新疆喀拉苏高等级墓地[24]、新疆石人子沟与西沟遗址[25]、秦始皇帝陵[26]等。本次研究发现血池北斗坊7 号坑的雄雌比为8:5,母马占有较高的比例,其背后蕴含的历史信息值得深思。另外,血池北斗坊7 号坑中出土葬马的毛色以栗色为主,其次为骝色,还有别的颜色,当时对于同一坑中马匹的毛色似乎没有严格地限定。
(2)运动能力/步态/肩高
中国古代的各个朝代都在有意识地培育身材高大、爆发力强、步态灵活的马种。为了适应长途骑乘,减少马背颠簸之苦,古人对驿马进行了选择,驯化可以走交替步法(溜蹄步)的马种,让骑马人感觉到舒适[27],例如,祁连山南北草原的大通马中,有大量天生会走这种步法的马;而东汉时期创作的“踏飞燕铜奔马”正是这种轻盈步态的反映。基于基因型和表型的关联性,血池北斗坊7 号坑中葬马的个体较小、不擅长交替步法、擅长长距离跑,似乎不能算严格意义上的良驹。

图一 北斗坊7 号坑与其他现代和古代家马的主成分散点图
四、结论
我们通过对陕西省雍山血池秦汉祭祀遗址北斗坊7 号坑中26 匹幼马马骨的古DNA分析,主要有两点认识:1.其母系来源的多样性极高,当时马匹的来源地可能不止一处,根据现代马和古代马的线粒体DNA 世系频率的主成分分析,血池北斗坊7号坑的样本更接近于现代的蒙古马,少数马的来源可能跟高海拔地区的马种有关。2.马匹中雄性略多、栗色马占大多数,当时对祭祀用马的性别、毛色的要求并不严格;这些马虽然耐力较好,但爆发力和步法灵活度一般,不太可能成长为出色的战马。这些认识为探讨当时祭祀用马的征集方式、选马标准等积累了重要的资料。今后我们还将继续对血池遗址其他祭祀坑中的用牲开展DNA 研究,在系列研究结果的基础上,努力破解历史长河中湮灭的千古之谜。
[1]Knapp M,Clarke A C,Horsburgh K A,et al.Setting the stage -building and working in an ancient DNA laboratory.[J].Annals of Anatomy,2012,194(1):3-6.
[2]Rohland N,Glocke I,Aximu-Petri A,et al.Extraction of highly degraded DNA from ancient bones,teeth and sediments for high-throughput sequencing [J].Nature Protocols,2018.
[3]Jansen,T.Mitochondrial DNA and the origins of the domestic horse [J].Proc Natl Acad Sci U S A,2002,99(16):10905-10910.
[4]赵若阳,赵一萍,李蓓,等.马毛色遗传机理研究进展[J].遗传,2018(05):21-32.
[5]Moon S,Lee J W,Shin D H,et al.A Genome-wide Scan for Selective Sweeps in Racing Horses [J].Asian Australasian Journal of Animal Sciences,2015,28(11):1525-1531.
[6]Hill E W,Jingjing G,Eivers S S,et al.A Sequence Polymorphism in MSTN Predicts Sprinting Ability and Racing Stamina in Thoroughbred Horses [J].PLoS ONE,2010,5(1):e8645.
[7]Andersson L S,Larhammar M,Memic F,et al.Mutations in DMRT3 affect locomotion in horses and spinal circuit function in mice [J].Nature,2012,488(7413):642-646.
[8]M Promerová,Andersson L S,Juras R,et al.Worldwide frequency distribution of the ‘Gait keeper’ mutation in the DMRT3 gene [J].Animal Genetics,2014,45(2):274-282.
[9]Fegraeus K J,Johansson L,Mäenpää M,et al.Different DMRT3 genotypes are best adapted for harness racing and riding in finnhorses [J].Journal of Heredity,2015,106(6):734-740.
[10]Makvandi-Nejad S,Hoffman G E,Allen J J,et al.Four Loci Explain 83% of Size Variation in the Horse [J].PLOS ONE,2012,7(7):e39929.
[11]Petersen J L,Mickelson J R,Rendahl A K,et al.Genome-Wide Analysis Reveals Selection for Important Traits in Domestic Horse Breeds [J].PLoS Genetics,2013,9(1):e1003211.
[12]Frischknecht M,Jagannathan V,Plattet P,et al.A Non-Synonymous HMGA2 Variant Decreases Height in Shetland Ponies and Other Small Horses [J].PLOS ONE,2015,10(10):1-11.
[13]Hendrickson S L.A genome wide study of genetic adaptation to high altitude in feral Andean Horses of the páramo [J].BMC Evolutionary Biology,2013,13(1):273-273.
[14]同[3].
[15]Cai D,Tang Z,Han L,et al.Ancient DNA provides new insights into the origin of the Chinese domestic horse [J].Journal of Archaeological Science,2009,36(3):835-842.
[16]蔡大伟,韩璐,朱泓.内蒙古赤峰地区青铜时代古马线粒体 DNA 分析[J].自然科学进展,2007(3).
[17]赵欣,李悦,陈洪海,等.陕西省淳化县枣树沟脑遗址马坑出土马骨的DNA 初步研究[J].南方文物,2015(3).
[18]赵欣,Rodrigues A T,尤悦,等.新疆石人子沟遗址出土家马的DNA 研究[J].第四纪研究,2014(1):187-195.
[19]赵欣,东晓玲,韩雨等.新疆木垒县平顶山墓群出土马骨的DNA 研究[J].南方文物,2017(3).
[20]蔡大伟,朱司祺,胡松梅,等.陕西凤翔秦公一号大墓车马坑马骨遗骸古DNA 研究[J].考古与文物,2018(3):106-112.
[21]Michael C,Melanie P,Norbert B,et al.Origin and History of Mitochondrial DNA Lineages in Domestic Horses [J].PLoS ONE,2010,5(12):e15311.
[22]同[20].
[23]Ruscillo D.Zooarchaeology:methods of collecting age and sex data [C]// In:Smith,C.(Ed.),Encyclopedia of Global Archaeology.Springer New York,New York,NY,2014.
[24]尤悦,于建军,陈相龙,等.早期铁器时代游牧人群用马策略初探—以新疆喀拉苏墓地M15 随葬马匹的动物考古学研究为例[J].西域研究,2017(04):108-120.
[25]李悦,尤悦,刘一婷,等.新疆石人子沟与西沟遗址出土马骨脊椎异常现象研究[J].考古,2016(1):108-120.
[26]陕西省考古研究所,秦始皇兵马俑博物馆.秦始皇帝陵园考古报告(2000)[M].北京:文物出版社,2006.
[27]甘肃农业大学.养马学[M].北京:农业出版社,1990.
猜你喜欢
临床肺科杂志(2022年3期)2022-11-26
中华医学图书情报杂志(2022年1期)2022-11-18
中华实用诊断与治疗杂志(2022年1期)2022-08-31
中国现代医生(2022年21期)2022-08-22
农村科学实验(2022年2期)2022-03-12
中国卒中杂志(2021年7期)2021-11-29
养猪(2021年4期)2021-08-26
三农资讯半月报(2020年2期)2020-03-09
中国体育科技(2018年6期)2018-12-13
小学阅读指南·低年级版(2017年10期)2017-11-20
