
冬凌草甲素抑制人宫颈癌HeLa细胞生存、迁移、侵袭的活性研究*
2020-04-12 03:18郭金兴
世界科学技术-中医药现代化 2020年9期
刘 艺,郭金兴
(1. 牡丹江医学院药学院 牡丹江 157011;2. 牡丹江医学院附属红旗医院 牡丹江 157011)
宫颈癌(cervical carcinoma)是常见的威胁女性健康及生命的恶性肿瘤。国际癌症研究中心研究数据表明,2013 年世界范围内确诊宫颈癌病例约48.5 万,死亡23.6 万[1]。我国宫颈癌统计学研究数据表明,2009 年,我国宫颈癌发病率为12.96/10 万,死亡率为3.28/10万[2]。宫颈癌95%的病例是由高危型人乳头瘤病毒(HPV)持续感染引起,其他危险因素如早婚、结婚次数过多、丈夫包皮过长、性伴侣过多、多产及初产年龄过早、人工流产次数过多、口服避孕药、吸烟与被动吸烟、机体免疫功能低下等[3-6]。目前宫颈癌的治疗仍以传统放化疗为主,而放化疗耐受已成为宫颈癌治疗失败、癌症转移和复发的主要原因之一[7]。近年来,中药治疗肿瘤以其多靶点、来源丰富、有效低毒的优势日益收到大多学者高度的重视。冬凌草甲素(oridonin)是唇形科香茶菜属植物冬凌草主要的抗癌有效成分,为贝壳杉烯类二萜。现代药理研究表明,冬凌草甲素对多种肿瘤有治疗作用如直肠癌[8]、前列腺癌[9]、乳腺癌[10]、肺癌[11]、肝癌[12]、食管癌[13]、卵巢癌[14]、胰腺癌[15]等,提示冬凌草甲素具有较好的抗肿瘤效果。也有研究表明,冬凌草甲素对人宫颈癌HeLa 细胞增殖具有明显的抑制作用[16,17]。然而有关于冬凌草甲素对HeLa细胞迁移和侵袭能力的影响鲜见报道。故此本研究观察了冬凌草甲素对HeLa 细胞迁移和侵袭能力及Wnt/β-连环蛋白(β-catenin)信号通路的影响,为其治疗宫颈癌提供实验依据。
1 材料与方法
1.1 实验材料
1.1.1 细胞株
人宫颈癌HeLa细胞(美国ATCC,货号:ZY-H066)接种于含10%热灭活胎牛血清(foetal bovine serum,FBS)、2%谷氨酰胺、100 μg·mL-1链霉素和100 U·mL-1青霉素的RPMI-1640 培养液中培养。每48 h 更换新鲜培养液,待细胞融合度达80%时,用0.25%的胰蛋白酶消化传代。
1.1.2 主要试剂
冬凌草甲素(纯度>99.4%,美国Amresco,货号:28957-04-2);二甲基亚砜(dimethyl sulphoxide,DMSO,美国 SIGMA,货号:D5879-100ML);RPMI-1640 培养基、FBS 和胰蛋白酶(美国 Gibco 公司,货号:12633、DXT-10099141、X4380);细胞计数试剂盒-8(cell counting kit-8,CCK-8);(美国 Abbkine 公司,货号:KTC011001);TUNEL 细胞凋亡原位检测试剂盒(美国Ambion 公司,货号:xyG001-1);Annexin V-FITC/PI 试剂盒(美国BD 公司,货号:556547);鼠抗人β-catenin单克隆抗体(美国Abcam 公司,货号:xyKF682);鼠抗人原癌基因(C-myc)单克隆抗体(美国Merck Millipore公司,货号:DXT-ST257261);鼠抗人细胞周期素D1(Cyclin D1)和兔抗人GAPDH 多克隆抗体(美国Bioss公司,货号:bs-0623R-1,bs-0755R-1);其它试剂均为进口或者国产分析纯。
1.1.3 主要仪器
Model 311 型CO2培养箱(美国Thermo Scientific 公司);Bio-Tek ELX800 型多功能酶标仪(美国Bio Rad公司);FACS CaliburTM流式细胞仪(美国BD 公司);BX43荧光显微镜(日本Olympus公司);Power PAC1000电泳仪(美国Bio Rad 公司);Micropulser 电转仪(美国Bio Rad公司)。
1.2 方法
1.2.1 药物配制
冬凌草甲素溶于 DMSO 配制成 160 μmol·L-1储备液,DMSO浓度≤0.1%且不诱导细胞增殖或者死亡。-20℃冰箱保存,临用时以RPMI-1640 培养液稀释到所需浓度。
1.2.2 CCK-8实验
严格按照CCK-8 检测试剂盒操作说明书进行HeLa细胞活力检测。将对数生长期HeLa细胞接种于96孔板,细胞密度为1.5× 104个细胞/孔。37℃、5%CO2及饱和湿度下培养,待细胞贴壁后,实验组分别加入含不同浓度为10、20、40、80、160 μmol·L-1冬凌草甲素的RPMI-1640培养液100 μL,对照组加入不含冬凌草甲素的RPMI-1640 培养液100 μL。每个剂量组均设6 个复孔。37℃、5% CO2及饱和湿度下培养。分别于12 h,24 h和36 h后终止培养,各孔均加入CCK-8试剂10 μL,混匀。37℃、5%CO2及饱和湿度下培养4 h。酶标仪检测450 nm 处的吸光度(optical density,OD)值,以未加入HeLa 细胞,未加入冬凌草甲素,但是加入CCK-8 试剂的培养空作为空白组。计算细胞活力,细胞活力(%)=(实验组的OD 值-空白组的OD 值)/对照组的OD 值-空白组的OD 值)× 100%,并应用Bliss 法计算半数抑制浓度(IC50)值。
1.2.3 细胞凋亡检测
(1)TUNEL染色
严格按照TUNEL 凋亡检测试剂盒操作说明书进行HeLa 细胞凋亡检测。取洁净无菌玻片置于24孔板底部,将对数生长期HeLa 细胞以密度为4 × 104个细胞/mL 接种于24 孔板。37℃、5%CO2及饱和湿度下培养,待细胞贴壁后,实验组分别加入含不同浓度为10、20、40 μmol·L-1冬凌草甲素的 RPMI-1640 培养液 300 μL,对照组加入不含冬凌草甲素的RPMI-1640培养液300 μL。每个剂量组均设6 个复孔。37℃、5% CO2及饱和湿度下培养24 h 后终止培养,取出玻片,4%多聚甲醛于4℃下固定 30 min,3% H2O2封闭 10 min,0.1%的 Triton-100 置于冰上打孔 5 min,加 80 μL 的 TUNEL检测液,37℃、5%CO2及饱和湿度下培养2 h,DAPI 染核5 min,50%甘油封片。荧光显微镜下观察HeLa 细胞核荧光变化。结果判定:凋亡细胞核发出绿色荧光,未凋亡细胞核发出蓝色荧光。随机选取10个视野观察,分别计数凋亡细胞数和总细胞数,计算凋亡指数。凋亡指数=凋亡细胞数/总细胞数×100%。
(2)Annexin V-FITC/PI双染法
严格按照AnnexinV-FITC/PI试剂盒操作说明书进行HeLa 细胞凋亡检测。分组及细胞培养同“(1)”。0.25%胰蛋白酶消化,1200 r·min-1离心5 min,收集细胞,PBS洗涤2次,250 μL结合缓冲液重悬细胞(Binding Buffer),2000 r·min-1离心 5 min 后,去除上清液,添加100 μL Binding Buffer 并吹打,依次加入 Annexin VFITC 和 PI 液各 5 μL 混匀,室温避光反应 15 min,再加入400 μL Binding Buffer,转移至流式管,于1 h 内流式细胞仪检测细胞凋亡,计算凋亡率。凋亡率(%)=Q2凋亡率+Q4凋亡率。
1.2.4 细胞粘附实验
将分组及细胞培养同“(1)”。预处理过的HeLa细胞接种于预先用Matrigel 胶包被96 孔板。37℃、5%CO2及饱和湿度培养120 min。PBS 洗涤2 次去除未粘附的细胞,加入MTT 20μL,37℃、5%CO2及饱和湿度培养4 h,加DMSO 150 μL 溶解结晶颗粒,置摇床上低速振荡10 min,酶标仪检测450 nm 处的OD 值,计算粘附率。粘附率(%)=实验组的OD/对照组的OD×100%。
1.2.5 划痕修复实验
将对数生长期HeLa 细胞以密度为4×104个细胞/mL接种于6孔板,37℃、5%CO2及饱和湿度下培养,待细胞平铺面积达80%后,用已消毒的10 μL 枪头垂直划一条泳道,PBS 洗去脱落细胞。分组及细胞培养同“(1)”。倒置显微镜下拍照,用NIH image J 软件计算划痕面积,计算迁移率。迁移率(%)=实验组划痕面积/对照组划痕面积×100%。
1.2.6 Transwell小室体外侵袭实验
将Transwell 侵袭小室进行Matrigel 胶铺胶,置于恒温培养箱中进行固化处理。用无血清培养液将HeLa 细胞饥饿培养24 h 后,在上层小室加入200 μL密度为4 × 104个细胞/mL 细胞悬液,而在下室中加入完全培养基。分组及细胞培养同“(1)”。取出小室,弃上层小室培养液,PBS 洗涤3 次,无菌棉签擦去未侵袭细胞,结晶紫染色,倒置显微镜下计数穿过的细胞总数,计算侵袭率。侵袭率(%)=实验组侵袭细胞数/对照组侵袭细胞数×100%。
1.2.7 Western blot分析
分组及细胞培养同“(1)”。1200 r·min-1离心 5 min,收集细胞,充分裂解后提取总蛋白,用BCA 法测定蛋白浓度。在10%十二烷基硫酸钠-聚丙烯酰胺预制凝胶中,各培养孔加入50 μg 蛋白样品进行电泳。常规转膜后5%脱脂奶粉封闭2 h。分别与Ⅰ抗鼠抗人β-catenin 单克隆抗体,鼠抗人C-myc 单克隆抗体,鼠抗人Cyclin D1 4℃孵育过夜,TBST洗膜3次,洗膜后加辣根过氧化酶标记山羊抗兔Ⅱ抗,37℃、5% CO2及饱和湿度下孵育1 h。TBST 洗膜3 次,ECL 显影。以GAPDH作为内参。Gel-pro 4.0软件进行分蛋白条带与GAPDH灰度比值,计算相对表达量。

图1 不同浓度冬凌草甲素对HeLa细胞增殖的影响
1.3 统计分析
采用SPSS 18.0 统计软件进行。数据分析计量资料采用“”表示,各组数据符合正态分布,方差齐时,采用单因素方差分析,组间两两比较,采用LDS 检验;方差不齐时采用 Games-Howell 检验。P <0.05 表示差异有统计学意义。
2 结果
2.1 冬凌草甲素对HeLa细胞增殖的影响
HeLa细胞经不同浓度0、10、20、40、80、160 μmol·L-1冬凌草甲素处理 12 h,24 h 和 36 h 后,以 CCK-8 检测细胞活力。结果显示,冬凌草甲素对HeLa 细胞增殖具有明显的抑制作用,且随着作用时间和作用浓度的增加,抑制作用逐渐增加(图1)。当冬凌草甲素浓度为 80 μmol·L-1处理24 h 时,HeLa 细胞活力约为 50%。24 h,48 h 和 72 h 的IC50分别为(151.36 ± 18.72)μmol·L-1,(62.10 ± 7.39)μmol·L-1和(8.41 ± 1.07)μmol·L-1。本研究后续实验采用 ≤80 μmol·L-(110、20、40 μmol·L-1)冬凌草甲素及24 h作为实验浓度和处理时间。
2.2 冬凌草甲素对HeLa细胞凋亡的影响
2.2.1 TUNEL染色结果
与对照组比较,10、20、40 μmol·L-1冬凌草甲素组HeLa 细胞凋亡指数显著升高(P< 0.05,P< 0.01)(图2、图3)。
2.2.2 Annexin V-FITC/PI双染法结果
与对照组比较,10、20、40 μmol·L-1冬凌草甲素组HeLa 细胞凋亡率显著升高(P< 0.05,P< 0.01)(图4、图5)。
2.3 冬凌草甲素对HeLa细胞粘附力的影响
与对照组比较,10、20、40 μmol·L-1冬凌草甲素组HeLa细胞粘附率显著降低(P< 0.05,P< 0.01)(图6)。
2.4 冬凌草甲素对HeLa细胞迁移能力的影响
与对照组比较,10、20、40 μmol·L-1冬凌草甲素组HeLa 细胞迁移率显著降低(P< 0.05,P< 0.01)(图7、图8)。
2.5 冬凌草甲素对HeLa细胞侵袭能力的影响
与对照组比较,10、20、40 μmol·L-1冬凌草甲素组HeLa 细胞侵袭率显著降低(P< 0.05,P< 0.01)(图9、图10)。
2.6 冬凌草甲素对HeLa细胞侵Wnt/β-catenin信号通的影响
与对照组比较,10、20、40 μmol·L-1冬凌草甲素组HeLa 细胞β-catenin、C-myc 和 Cyclin D1 蛋白表达均显著降低(P< 0.05,P< 0.01)(图11、图12)。
3 讨论
本研究CCK-8 结果显示,冬凌草甲素可呈剂量-时间依赖效应的方式抑制HeLa 细胞增殖。同时本研究TUNEL 染色和Annexin V-FITC/PI 双染法结果显示,冬凌草甲素亦可诱导HeLa 细胞凋亡。以上研究结果与文献报道相一致[18]。
迁移和侵袭是恶性肿瘤转移和浸润的重要过程,迁移和侵袭能力越强说明肿瘤细胞运动能力越强,肿瘤越容易转移和浸润[19]。转移和浸润是一个受多种因素调控的复杂过程,除了与肿瘤细胞新生血管的形成有关以外,亦与细胞的粘附力,迁移和侵袭能力的改变有关,为恶性肿瘤治疗失败及预后较差的主要原因[20]。本研究通过细胞粘附实验检测了冬凌草甲素对HeLa细胞粘附力的影响,划痕修复实验检测了冬凌草甲素对HeLa细胞侵袭能力的影响,Transwell小室体外侵袭实验检测了冬凌草甲素对HeLa细胞侵袭能力的影响。结果显示,冬凌草甲素显著抑制HeLa 细胞粘附力,迁移和侵袭能力,且随着药物浓度增加,抑制作用逐渐增加。这些结果提示,冬凌草甲素除了可以抑制HeLa细胞增殖和诱导其凋亡以外,亦可通过抑制其迁移和侵袭能力共同发挥抗宫颈癌作用。
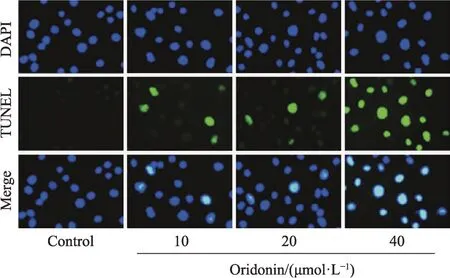
图2 TUNEL染色结果(×200)

图3 冬凌草甲素对HeLa细胞凋亡指数的影响

图4 Annexin V-FITC/PI双染法结果

图5 冬凌草甲素对HeLa细胞凋亡率的影响

图6 冬凌草甲素对HeLa细胞粘附率的影响

图7 划痕修复实验结果(×40)

图8 冬凌草甲素对HeLa细胞迁率的影响

图9 Transwell小室体外侵袭实验结果(结晶紫,×200)

图10 冬凌草甲素对HeLa细胞侵袭率的影响
宫颈癌细胞迁移和侵袭是制约其疗效、影响预后及的重要因素之一。恶性肿瘤细胞的转移和浸润是一个由多种分子参与调节的复杂过程。Wnt/β-catenin信号通路是细胞内重要信号通路,与恶性肿瘤有着密切的关系,参与多个生物学进程调控(如调控细胞的增殖、凋亡、迁移、侵袭等),其异常活化参与人类多种癌症的发病过程[21]。β-catenin 是 Wnt/β-catenin 信号通路关键的信号转导分子,是该信号通路激活的标志[22]。β-catenin表达量越高,肿瘤的恶性程度越高,迁移和侵袭能力越强,预后也越差[23]。当Wnt/β-catenin信号通路异常激活时,β-catenin 在细胞质中降解减少,去磷酸化并进入细胞核,进而开启下游C-myc 和Cyclin D1 等靶基因的异常表达,从而促进肿瘤细胞的增殖,转移和侵袭[24]。研究表明,宫颈癌患者及HeLa细胞β-catenin 表达异常[25,26]。本研究 western blot 分析结果显示,冬凌草甲素显著降低HeLa 细胞β-catenin、C-myc 和Cyclin D1 表达。以上结果提示,冬凌草甲素可抑制Wnt/β-catenin信号通路。

图11 Western blot结果
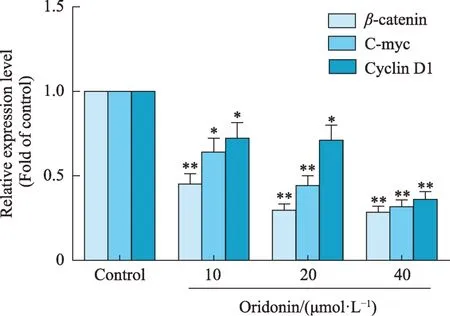
图12 冬凌草甲素对HeLa细胞Wnt/β-catenin信号通的影响
综上所述,冬凌草甲素具有抑制HeLa 细胞增殖的作用,亦可诱导凋亡及抑制迁移和侵袭能力,并抑制Wnt/β-catenin 信号通路。目前对于冬凌草甲素的抗宫颈癌作用仍不明确,今后的实验中还需要具体探讨。
猜你喜欢
中华实用诊断与治疗杂志(2022年1期)2022-08-31
中华实用诊断与治疗杂志(2022年1期)2022-08-31
中学生物学(2021年8期)2021-11-02
疯狂英语·新策略(2021年7期)2021-08-26
服饰导报·鞋世界(2021年4期)2021-05-17
山西农业大学学报(自然科学版)(2020年1期)2020-03-04
绿色科技(2019年18期)2019-11-22
药学研究(2015年11期)2015-12-19
科学启蒙(2015年8期)2015-08-07
中国民族民间医药·下半月(2011年4期)2011-09-27
