
Microbial communities present on mooring chain steels with diff erent copper contents and corrosion rates*
2020-03-19 12:30WUJiajiaGAOJieyanZHANGDunTANFaqiYINJiangWANGYuSUNYanLIEe
WU Jiajia GAO Jieyan ZHANG Dun TAN Faqi YIN Jiang WANG Yu SUN Yan LI Ee
1 Key Laboratory of Marine Environmental Corrosion and Bio-fouling, Institute of Oceanology, Chinese Academy of Sciences,Qingdao 266071, China
2 Open Studio for Marine Corrosion and Protection, Qingdao National Laboratory for Marine Science and Technology, Qingdao 266237, China
3 Center for Ocean Mega-Science, Chinese Academy of Sciences, Qingdao 266071, China
4 University of Chinese Academy of Sciences, Beijing 100049, China
5 Shanghai Bainite Chain Material Tech Co. Ltd., Shanghai 200439, China
Abstract Copper has long been utilized as a disinfectant for bacteria, but its impact on microbial communities attached to the steel surface in seawater remains unknown. In the present study, 3 mooring chain steels of diff erent copper contents are subjected to a 3-month marine f ield exposure, and the corrosion rate increases in the order of BR5 steel (without copper) < BR5CuH steel (0.8% copper) < BR5CuL steel(0.4% copper). The microbial community results show that copper introduction does not result in an obvious change in microbial quantity, but it alters the diversity, richness, and structure of microbial communities due to the variation in copper-resistance of diff erent species. BR5CuH steel holds microbial communities with the highest percentage of some well-known corrosive microbes including sulfate-reducing bacteria, sulfuroxidizing bacteria, and iron-oxidizing bacteria, but possesses the lowest community diversity/richness owing to the toxicity of copper. The microbial community diversity/richness is stimulated by the low-copper content of BR5CuL steel, and this steel also carries an intermediate proportion of such corrosive bacteria.Both well-known corrosive bacteria and microbial community diversity/richness seem to be involved in the corrosion acceleration of copper-bearing mooring chain steels.
Keyword: marine corrosion; microbially inf luenced corrosion; microbial community; mooring chain steel;copper introduction
1 INTRODUCTION
Seawater is a rather harsh electrolyte for metal corrosion. Additional to the high salt concentration,diverse microorganisms survived play quite an important role in the corrosivity. Corrosion aff ected by microorganisms is known as microbially inf luenced corrosion (MIC), a destructive type of corrosion.Approximately 20%-30% of all corrosion failure is related to MIC, with a direct f inancial cost of $30-50 billion per year (Javaherdashti, 2008). There are numerous cases for the marine corrosion deterioration due to MIC like a rapid perforation of welded stainless steel pipes in a yacht served in Auckland with a corrosion rate of 40 mm/a (Liu, 2014), accelerated low water corrosion of steel pilings in some harbours around the world (Melchers and Jeff rey, 2013), and severe pitting corrosion of mooring chains served in West Africa (Fontaine et al., 2012). MIC is normally achieved by the biof ilm from the attachment of microorganisms onto the substrate, and two kinds of mechanisms are involved in metal materials, i.e.,extracellular electron transfer-MIC and metabolite-MIC (Xu et al., 2016; Li et al, 2018; Gu et al., 2019;Jia et al., 2019). It is expected to be suppressed via the inhibition on biof ilm formation and development.Several methods have been proposed and utilized for biof ilm inhibition, which can be divided into four categories: modif ication in chemical compositions and structures of the substrate metals, surface modif ication with coatings or functional f ilms, cathodic protection,and introduction of bactericides into bulk solutions.Among them, substrate metal modif ication by the addition of alloying elements has attracted extensive attention due to its simplicity and f lexibility.
Copper is a common alloying element in steel manufacture to promote mechanical performance (Kim et al., 2011), and its addition can also improve the atmospheric corrosion resistance of high strength low alloy (HSLA) steels owing to the formation of adherent and protective rust layers (Lins et al., 2017). A typical copper-bearing HSLA steel is ASTM A690 steel.Because copper has long been utilized as a disinfectant for bacteria, the introduction of copper into steels might favor their resistance against MIC. There are extensive reports on MIC inhibition by copper addition into stainless steels, especially the work of Xu and Yang et al. Copper has been added into stainless steels with the designations like 304, 304L, 317L, and 2205 to resist corrosion fromEscherichiacoli,Staphylococcus aureus, andPseudomonasaeruginosa, etc. (Hong and Koo, 2005; Nan et al., 2015; Xia et al., 2015; Sun et al.,2016). The MIC inhibition of copper-bearing stainless steels is closely related to the release of copper ions,which kill bacteria via damaging cytoderm and cytomembrane, generating the reactive oxygen species,and other mechanisms (Espírito et al., 2011; Warnes et al., 2012; Sun et al., 2016). Although these reports are signif icant for the development of copper-bearing steels, two points ought to be stressed before their practical applications in seawater.
Firstly, how about the impact of copper introduction on MIC of HSLA steels? Unlike the case for stainless steels, there are hardly any reports on HSLA steels that are widely utilized in marine structural construction, except for the very recent work by Shi et al. (2018). They found that the as-aged copper-bearing X80 pipeline steel (1.06% copper) with Cu-rich precipitates inhibited the attachment of sulfatereducing bacteria (SRB) andPseudomonas aeruginosa, and therefore had better corrosion resistance than bare X80 steel. Consequently, further studies are highly desirable on this topic.
Secondly, how copper introduction aff ects microbial communities and what is the relationship between microbial communities and corrosion of copper-bearing steels in seawater? Normally, the effi ciency of copper-bearing steels against MIC was evaluated with specif ic isolated bacterial strains in laboratories, and to the best of our knowledge, there has been no reports focused on the impact of copper introduction on microbial communities in seawater.Microbial communities seem to be sensitive to the variation in material surface features and the environments, and they vary with substrate material types, surface microscale topography of organic f ilms,organic biocides in antifouling coatings, chemical composition of solutions, and environmental factors like temperature. It is expected that microbial communities will be aff ected by copper introduction due to its release from the steel surface, but the accurate changes need further investigation. Although there are several reports on the impact of copper addition on the corrosion rate of HSLA alloys in seawater by f ield experiments, the role of copper is still in controversy: some reports demonstrate that it is strongly benef icial (Petersen, 1977), while others indicate deleterious or no eff ects (Forgeson et al.,1960; Hou et al., 2000). These conf licts are closely related to the diff erence in copper contents, exposure time, and seawater features. Melchers (2003, 2004)gave a new comparative analysis of these reported contradictory data for copper-bearing steels based on a multi-phase phenomenological corrosion-time model, and proposed that copper had relatively little eff ect on corrosion under aerobic conditions, inhibited the development of anaerobic corrosion, and increased the rate in the fully developed anaerobic corrosion.Although he did not give much explanation for the diff erent impacts of copper at diff erent stages, SRB were assumed to have a greater inf luence on the corrosion behavior of copper-bearing steels than that of copper-free steels. Unfortunately, no eff orts have been made to analyze the inf luence of copper addition on microbial communities, which is in urgent need for the comprehension of corrosion of copper-bearing steels in seawater.
In this study, mooring chain steels, typical HSLA steels, were selected as research subjects, and the eff ect of copper addition on corrosion rate in seawater was investigated by f ield experiments. Furthermore,microbial communities on diff erent steels were investigated with the aid of f luorescence microscope observation and high-throughput Illumina Miseq sequencing, and the relationship between bacterial communities and corrosion rate of steels was analyzed.The present work is meaningful for the comprehension on the corrosion mechanism of copper-bearing HSLA steels and their potential applications in seawater.


Fig.1 Metallographic microstructures of different steels observed by an optical microscopy
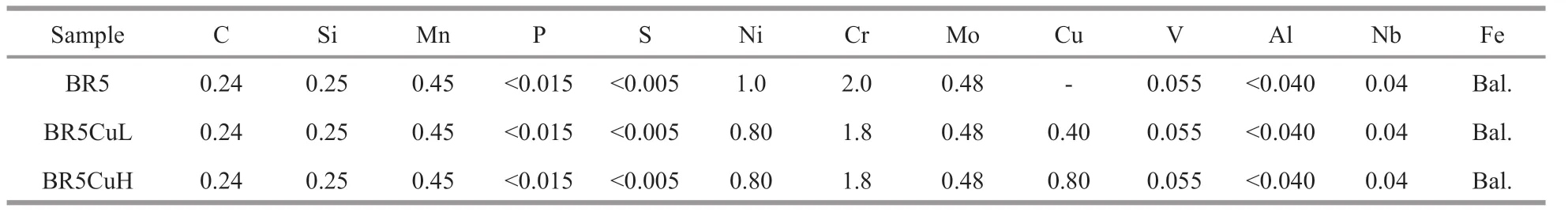
Table 1 Chemical compositions of diff erent steels (in wt%)
2 MATERIAL AND METHOD
2.1 Steel coupon preparation and sampling
Diff erent amounts of high purity copper were added together with other raw materials during melting to give mooring chain steels with diff erent copper contents. The chemical compositions of diff erent steels were analyzed by the optical emission spectroscopy method, and are reported in Table 1. All steels gave a phase structure of composite bainite(Fig.1), and copper introduction had little impact on the grain size, which was around 6.7 μm measured by employing the linear intercept procedure according to ASTM E112. Flat tensile specimens were fabricated according to SATM D3039-76, and the copper alloyingled to a slight enhancement in strength (Table 2).
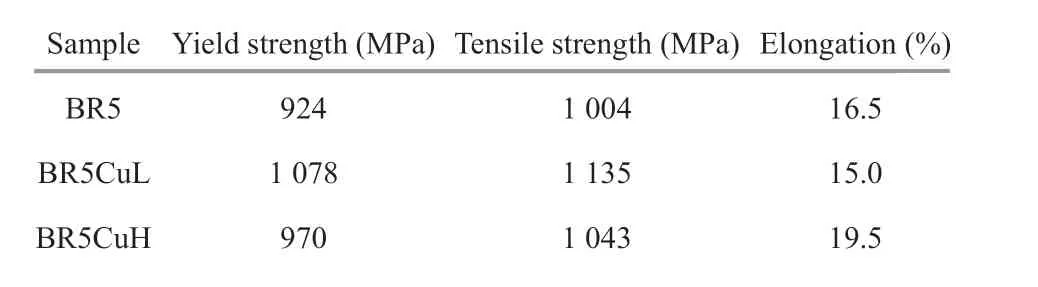
Table 2 Room-temperature tensile test results
Coupons with the size of 200 mm× 100 mm× 2 mm were cut from steel plates, abraded sequentially with a series of silicon carbide papers with grit sizes of 400, 600, and 800, cleaned by ethanol in an ultrasonication bath, dried by nitrogen, and weighed with a microbalance. Coupons were f ixed to a coated frame and hung about 1 m below the lowest tide level at a seawater corrosion station in Qingdao, China(36°03′N; 120°25′E) from 27 July 2017 to 27 October 2017. The average seawater temperature, dissolved oxygen concentration, pH, and salinity during this period were 22.1°C, 8.3 mg/L, 8.2, and 31%, respectively.Three duplicates were done for each steel.
At the end of the immersion experiments, all samples were removed from seawater, photographed,carried to the laboratory in sealed sterile Whirl-Pak bags within 1 h, and processed immediately. Corrosion product layers were scraped randomly at diff erent sites of all the corroded coupons, placed into two beakers holding 50 mL seawater sterilized via autoclaving for each steel, and weighed. The weight of corrosion products (Δm) in each beaker was obtained from the diff erence between values before and after product introduction. The beakers were shaken vigorously to give uniform suspensions,which were then subjected to DNA extraction. Half of the suspension was f ixed with 0.1% formaldehyde for bacterial cell enumeration later.
2.2 DNA extraction, amplication, sequencing, and data processing
Corrosion product suspensions for each steel in diff erent beakers were utilized as independent duplicates for DNA extraction after f iltration with 0.22 μm membrane, which was carried out with a Power Soil DNA Extraction Kit (MO BIO, Carlsbad,CA, USA) according to the manufacturer’s instructions. The purity of the DNA extracts was checked using a Nanodrop ND-1000 spectrophotometer(Nanodrop Thermo Scientif ic, Wilmington, DE,USA), and quantif ied with a Qubit 2.0 f luorimeter(Qubit, Invitrogen, Carlsbad, CA, USA) using the dsDNA BR Assay Kit (Invitrogen) according to the manual.
The V3-V4 region of 16S rRNA gene was amplif ied by polymerase chain reaction (PCR) with the primer pair 338F (5′-ACTCCTACGGGAGGCAGCAG-3′)and 806R (5′-GGACTACHVGGGTWTCTAAT-3′)following the protocol described by Li et al. (2017).After amplif ication, PCR products were analyzed on 2% agarose gel via electrophoresis to determine the relative intensity of bands, and then purif ied using the AxyPrep DNA Gel Extraction Kit (Axygen Biosciences, Union City, CA, USA). The purif ied amplicons were subjected to quantitative PCR using Quant-iTPicoGreen dsDNA Assay Kit (Invitrogen) at a microplate reader (BioTek, FLx800), and pooled accordingly. The f inal PCR products were used to prepare the DNA library by following the Illumina TruSeq DNA library preparation protocol, and then paired-end Illumina MiSeq sequencing was performed.
Raw sequences were demultiplexed and qualityf iltered using the default parameters in QIIME software package to obtain high-quality clean tags.Chimeras were detected and removed using the de novo mode of UCHIME, and the remaining sequences were subsequently clustered into operational taxonomical units (OTUs) at 97% similarity.Representative sequences with the highest abundance were chosen from each OTU, and aligned against the Greengenes database using PyNAST.
A Venn diagram was created using Mothur v.1.30.1 to identify the common and unique OTUs among microbial communities formed on diff erent steels.Rarefaction curves were plotted based on the aligned OTU table using QIIME, and then alpha diversity analysis including Chao1, ACE, Shannon, and Simpson indexes was evaluated. The relative abundance of OTUs at diff erent taxonomic levels was plotted as bar graphs with the aid of R program, and a heatmap at the genus level was also drawn.
2.3 Microbial cell enumeration
The corrosion product suspension f ixed by formaldehyde was stained with acridine orange with a f inal concentration of 0.1% for 5 min in the dark, and then 1 mL suspension was syringe f iltered through Whatman polycarbonate f ilters (pore size: 0.2 μm)previously stained with Sundan Blank. The f ilters were mounted on microscopic glass slides, and examined under a f luorescence microscope (Axio Imager A2; Carl Zeiss Microimaging GmbH; Jena,Germany). For each f ilter, at least 15 f ields were selected and counted to give an average value (Na).The cell number per g of corrosion products was 50NaS/(SfΔm), in whichSandSfwere the area of f ield and f ilter, respectively.
2.4 Corrosion product characterization, weight loss measurement, and morphology observation
Corrosion products scraped were subjected to X-ray diff raction (XRD) for chemical composition analysis. XRD spectra were recorded with Cu Kα radiation at 40 kV and 40 mA, and scans were stepped from 10° to 80° at a rate of 10°/min. After the surface was scrubbed carefully with a brush, the coupons were cleaned with the Clark’s solution containing 20 g antimony trioxide and 50 g stannous chloride in 1 000 mL concentrated hydrochloric acid (ASTM G1-03). Then they were rinsed with distilled water,cleaned in absolute ethanol, dried with nitrogen, and weighed. Corrosion rate in the unit of μm/a was calculated from the weight loss divided by the product of steel density, exposure time, and surface area.Corrosion morphology after corrosion product removal was recorded with a digital camera.

Fig.2 Digital photos for diff erent steels before (a) and after the removal of corrosion products (b-d, b: BR5, c: BR5CuL, d:BR5CuH)
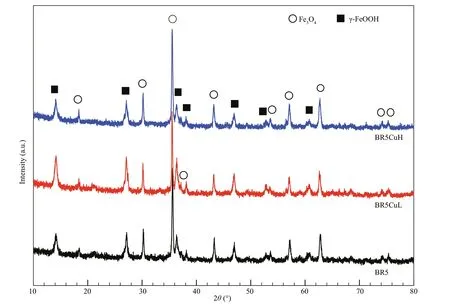
Fig.3 XRD spectra recorded on corrosion products of diff erent steels
3 RESULT
3.1 Corrosion morphology, product, and rate
After 3 months of f ield exposure in seawater, the surface of all steel coupons was fully covered by corrosion products with the color of brown and brick red (Fig.2a). When the products were removed, it could be observed that samples corroded almost evenly with several sporadic shallow pits (Fig.2b-d).XRD results shown in Fig.3 demonstrated that all these steels gave similar corrosion products, mainly consisting of Fe3O4and γ-FeOOH, which are commonin f ield seawater immersion experiments. The calculated corrosion rate of diff erent steels is shown in Table 3, which increased in the order of BR5 steel< BR5CuH steel < BR5CuL steel with the values of 0.18, 0.34, and 0.24 mm/a, respectively. The introduction of copper promoted corrosion, and the promotion effi ciency was content-dependent, which was higher with the content of 0.4%.
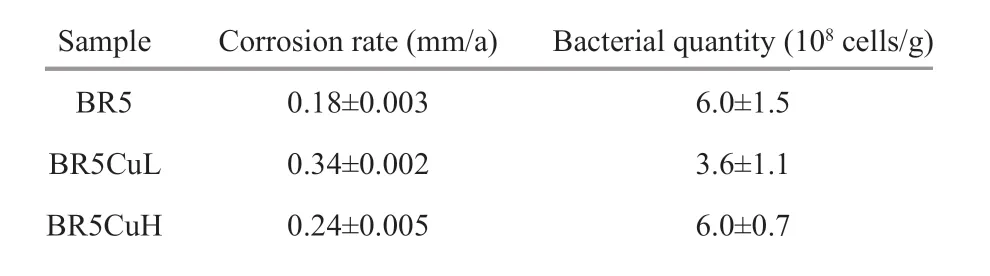
Table 3 Corrosion rate of diff erent steels and the microbial quantity of communities on them
3.2 Microbial counts
Corrosion products formed on diff erent steels gave similar microbial quantity with the magnitude of 108cells/g (Table 3), which was much higher than the values determined by methods based on cultivation(Yang et al., 2009). Diff erent from the case for single strains likePseudomonasaeruginosaandStaphylococcusaureus(Sun et al., 2016; Xu et al.,2018), the introduction of copper did not lead to an obvious decrease in the quantity of attached microbial cells for natural consortia in the present study. The antibacterial activity of copper may be considered carefully when copper-bearing HSLA steels are utilized in natural environments.
3.3 Diversity, richness, and composition of microbial communities
The raw paired-end reads obtained by MiSeq highthroughput sequencing were f iltered to remove potential erroneous sequences. 32 453 and 38 966 eff ective tags were yielded for the independent duplicates of BR5 steel, giving an average of 35 710.The average counts of eff ective tags for BR5CuL and BR5CuH steels were 38 367 and 38 813. Tags were grouped into OTUs at a 97% similarity level, and an average of 818 OTUs was gained from BR5 steel duplicates with the numbers of 812 and 824. The average number of OTUs for BR5CuL and BR5CuH steels was 878 and 631 (Table 4), respectively.Therefore, microbial community diversity increased with the addition of 0.4% copper, and decreased with a further increase in copper content to 0.8%.
All rarefaction curves exhibited in Fig.4 tended toapproach the saturation plateau, indicating the sequencing data are suffi cient to assess the complete population. The highest OTU richness was observed on BR5CuL steel, and the lowest was on BR5CuH steel. Furthermore, alpha diversity indexes were calculated and displayed in Table 4, in which Chao1 and ACE indices ref lected the community richness,while Shannon and Simpson indexes were related to the community diversity (Liu et al., 2015b). The higher these indices, the more community diversity,and richness had. Consequently, microbial communities formed on BR5CuL steel possessed the highest diversity and richness, and those on BR5CuH steel gave the lowest. Similar to the case of corrosion rate, the impact of copper introduction on community diversity and richness was also dependent on its contents, in which 0.4% was benef icial and 0.8% was detrimental.
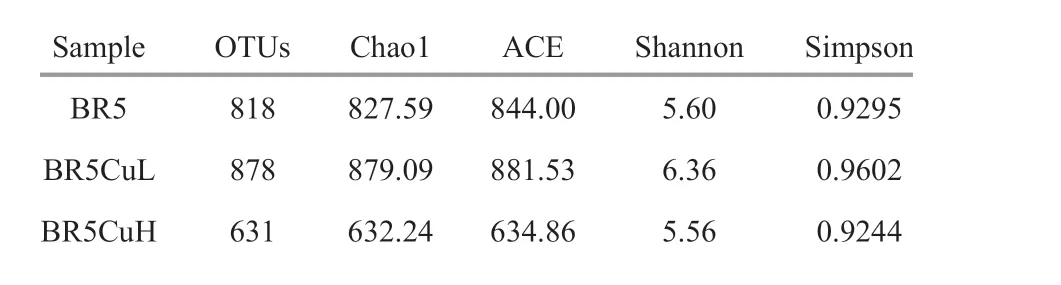
Table 4 OTU numbers and alpha diversity estimates for microbial communities formed on diff erent steels
The Venn diagram was constructed to compare common and unique OTUs among diff erent steels,and the result is displayed in Fig.5. Total 1 184 OTUs were presented for microbial communities formed on BR5 steel, which was derived from the summation of 812 and 824 for individual duplicates with the subtraction of 452 common OTUs. Similarly, there were total 1 260 and 895 OTUs for communities on BR5CuL and BR5CuH steels, respectively. All microbial communities on 3 steels carried a total of 1 980 OTUs, but only 431 of them were shared by all steels, accounting for 21.8% of the total OTUs. The shared OTUs covered 12 phyla, and Proteobacteria dominated with the proportion of 83.1%, which is followed by Firmicutes (6.3%), Actinobacteria(3.2%), and Bacteroidetes (2.8%). The number of unique OTUs was 418, 415, and 219 for BR5,BR5CuL, and BR5CuH steels, respectively,accounting for 53.1% of the total OTUs. Therefore,the variation in microbial community composition was remarkable among diff erent steels. Furthermore,this variation between BR5 and BR5CuH steels was larger evidenced by the lower ratio of common OTUs(32.8%) than that for BR5CuL-BR5 (38.8%) and BR5CuL-BR5CuH (37.8%) couples.
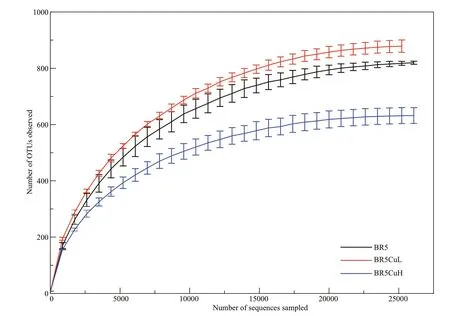
Fig.4 Rarefaction curves of the OTU number for microbial communities formed on diff erent steels
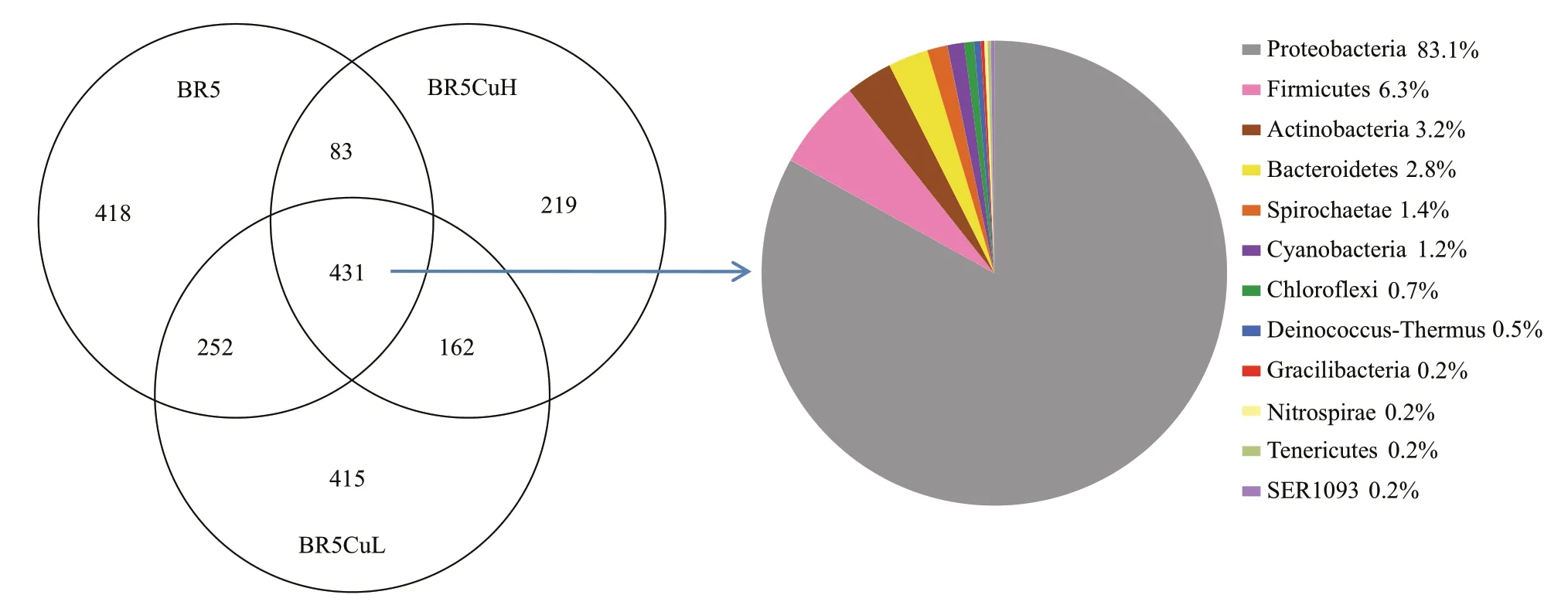
Fig.5 Venn diagram showing the overlap of OTUs between microbial communities formed on diff erent steels, and the taxonomic identity of the shared OTUs at phylum
The composition of bacterial communities was further analyzed at diff erent taxonomic levels, and the results are displayed in Fig.6. At the phylum level,Proteobacteria dominated in microbial communities formed on all steels with a proportion of more than 90%, which was in good accordance with its predominance in shared OTUs exhibited in Fig.5.Proteobacteria is a major phylum of gram-negative bacteria, and ubiquitous in marine biof ilms (Salta et al., 2013; McBeth and Emerson, 2016). The f ive most abundant classes of microbial communities all belonged to the phylum Proteobacteria, but their contents varied among diff erent steels. Class Gammaproteobacteria took a commanding leading in microbial communities formed on BR5 steel with the content of 62.9%, which was followed by Epsilon-(11.4%), Alpha- (7.6%), and Betaproteobacteria(6.8%). Superiority of Gammaproteobacteria was decreased on BR5CuL steel, and the proportion of Gamma-, Delta-, Alpha-, Epsilon-, and Betaproteobacteria was 40.8%, 27.8%, 9.2%, 7.8%,and 5.0%, respectively. For microbial communities formed on BR5CuH steel, it was Deltaproteobacteria(37.1%) that held a narrow lead over Gammaproteobacteria (35.7%), followed by Epsilon-(12.6%), Alpha- (6.0%), and Betaproteobacteria(4.4%). It was obvious that the proportion of Deltaproteobacteria increased with copper contents.
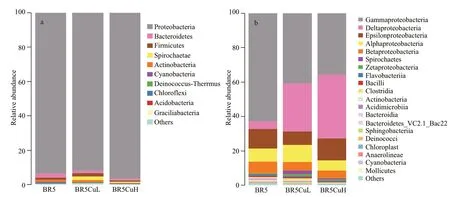
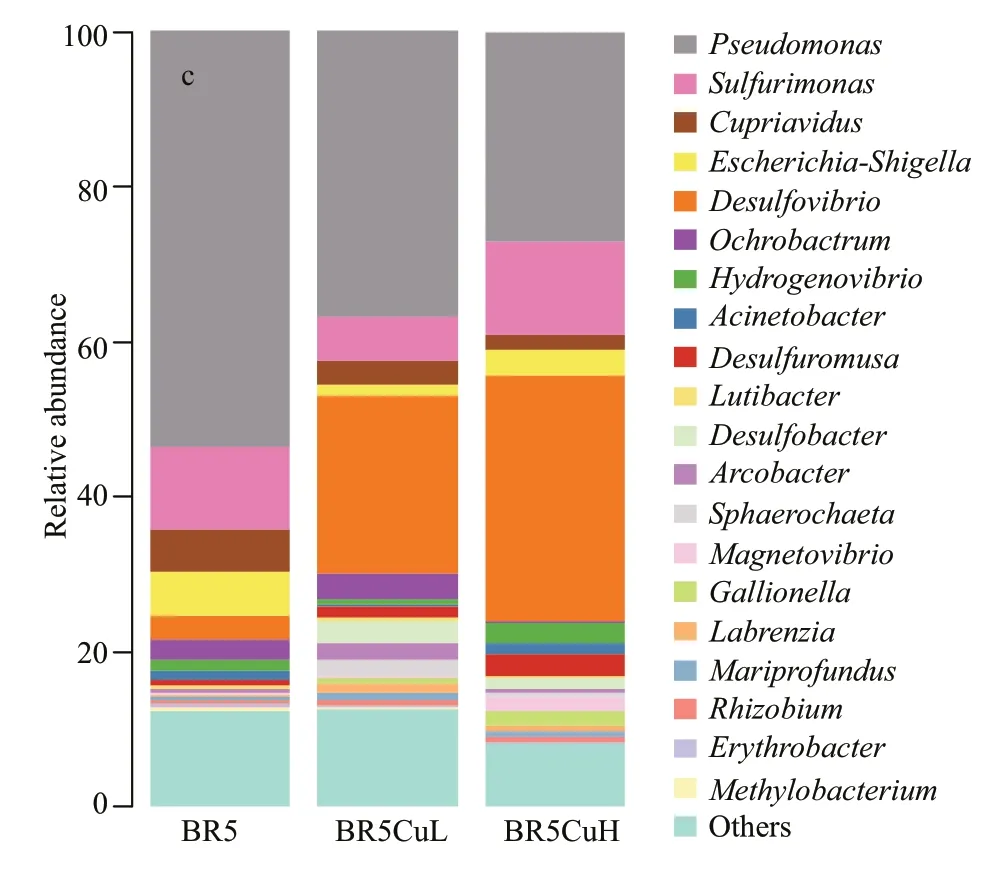
Fig.6 Relative abundance of microbial 16S rRNA gene sequences from communities on different steels presented at the phylum (a), class(b), and genus (c) levels
Figure 6c shows the proportion of the 20 most abundant genera in microbial communities on diff erent steels, and there were f ive genera of which the proportion reduced with an increase in copper contents, i.e.,Pseudomonas,Cupriavidus,Lutibacter,Erythrobacter, andMethylobacterium.Pseudomonasfrom the class Gammaproteobacteria dominated in biof ilms formed on BR5 and BR5CuL steels with a percentage of 53.4% and 36.8%, and it was the second most abundant genus for BR5CuH steel just behindDesulfovibrio(27.0% vs. 31.5%). Species and strains ofPseudomonasare normally aerobic microbes, and a signif icant number of them produce extracellular polymeric substances (EPS) to favor biof ilm formation, which are expected to aff ect metal corrosion. Corrosion of stainless steels is commonly enhanced byPseudomonasspecies via mechanisms like diff erential aeration cells, the chemical reaction between EPS and steel substrates, the production of siderophore, and the formation of CrN and CrO3(Abdolahi et al., 2014; Xu et al., 2017). By contrast,for carbon steel and low alloy steel materials that do not form a passive f ilm on the surface, the presence ofPseudomonassp. usually inhibits corrosion due to the decreased oxygen concentration by metabolism and the formation of compact biof ilms (Jayaraman et al.,1998; Volkland et al., 2000). The compact f ilms can prevent diff usion of corrosive species such as oxygen to the metal surface, thereby reducing the corrosion rate. Under extreme conditions without oxygen, some species likePseudomonasaeruginosapromote carbon steel corrosion via biocatalytic electron transfer when they survive with nitrate as the terminal electron acceptor (Jia et al., 2017). Since nitrate concentration was low in the present seawater(<2 μmol/L), such kind of corrosion promotion byPseudomonaswas rather limited.Cupriavidusaffi liated to class Betaproteobacteria occupied 5.6%,3.1%, and 1.8% of the bacterial communities on BR5,BR5CuL, and BR5CuH steels. Contents of the other three genera were no more than 0.6%, and the diff erence among diff erent steels was smaller than 0.4%. Species and strains of these four genera are obligate aerobic microbes, and not involved in sulfur or iron cycling that is believed to play an important role in corrosion impact from microbes. So far, there has been no knowledge on the corrosion of HSLA steels aff ected by specif ic strains of these genera.Consequently, their contribution to corrosion variation among diff erent mooring chain steels may be quite limited in the present study.
The percentage ofSulfurimonas,Escherichia-Shigella,Hydrogenovibrio, andAcinetobacterdecreased f irstly and then increased with the increase in copper contents. The ratio forSulfurimonaswas 10.6%, 5.6%, and 12.2% among diff erent steels, and the corresponding values forHydrogenovibriowere 1.3%, 0.5%, and 2.5%. As an Epsilonproteobacteria,Sulfurimonasis known for reducing nitrate, oxidizing both sulfur and hydrogen, which has been reported as a signif icant genus in off shore water injection systems treated with nitrate (Bødtker et al., 2008).Sulfurimonasoxidizes corrosive sulf ide generated by SRB to inhibit corrosion, but the produced sulfuric acid is also corrosive. Consequently, there have been some conf licts in the eff ectiveness of nitrate treatment,which are closely related to the specif ic environments(Nemati et al., 2001; Okoro et al., 2014). When reductive sulfur-containing compounds other than sulf ide are utilized as electron donors, the corrosivity ofSulfurimonasis expected due to corrosive sulfuric acid produced (Huber et al., 2016). Similar toSulfurimonas,Hydrogenovibriofrom class Gammaproteobacteria is also capable of oxidizing reduced sulfur compounds and hydrogen, and similar corrosion impact is expected. Gammaproteobacterial genusEscherichia-Shigellepossessed a proportion of 5.5%, 1.3%, and 3.4% for microbial communities on BR5, BR5CuL, and BR5CuH steels, and another Gammaproteobacterial genusAcinetobactergave values of 1.2%, 0.4%, and 1.5%. Their inf luence on steel corrosion is similar to other plain aerobic microorganisms likePseudomonas(Jayaraman et al., 1998).
The proportion of generaDesulfovibrio,Desulfuromusa,Magnetovibrio, andGallionellaimproved with copper contents.Desulfovibrio, a member of class Deltaproteobacteria, is a typical genus for SRB. Its content increased sharply from 3.2% to 23.1% with the addition of 0.4% copper into BR5 steel, and rose furthered to 31.5% with 0.8%copper. SRB have for long been viewed as the major corrosion-causing microbes in anoxic environments,and they constitute half of the total MIC loss (Chen et al., 2014). Extensive attention has been paid to steel corrosion aff ected by SRB, and several corrosion acceleration mechanisms have been proposed including cathodic depolarization by hydrogenase and iron sulf ide (Von Wolzogen Kühr and Van der Vlugt, 1934; King et al., 1973), production of phosphorous compounds (Iverson, 1998), direct electron transfer from Fe0(Dinh et al., 2004), and biocatalytic cathodic electron transfer (Li et al.,2015). The percentage of another Deltaproteobacterial genusDesulfuromusawas improved from 0.6% to 1.3% and 2.7% by copper addition. Species ofDesulfuromusacannot utilize sulfate as an electron acceptor, and they reduce elementary sulfur to sulf ide.Although there have been none work on corrosion mechanisms aff ected byDesulfuromusaspecies, their role in corrosion enhancement is expected from the corrosive sulf ide (Mand et al., 2016). The proportion ofMagnetovibrioandGallionellawas both 0.1% in bacterial communities formed on BR5 steel, increased to 0.2% and 0.8% on BR5CuL steel, and then rose further to 1.9% and 1.8% on BR5CuH steel,respectively. Species and strains ofMagnetovibrioincluded in class Alphaproteobacteria assimilate inorganic carbon chemolithoautotrophically with thiosulfate and sulf ide as the electron donors, and are typical sulfur-oxidizing bacteria (SOB). Furthermore,they synthesize magnetosomes containing magnetite crystals via biomineralization (Bazylinski et al.,2013).Magnetovibriocan accelerate metal corrosion by the corrosive sulfuric acid produced similar to SOB generaSulfurimonasandHydrogenovibriomentioned above.Gallionella, a member of class Betaproteobacteria, is a typical genus for ironoxidizing bacteria (IOB) that promote steel corrosion by accelerated transformation of Fe2+to Fe3+(Liu et al., 2015a).
There are another seven genera of which the proportion increased f irstly and then decreased with the increase in copper contents, and they gave the highest values in microbial communities formed on BR5CuL steel.Ochrobactrumfrom the class Alphaproteobacteria possessed a percentage of 2.6%,3.3%, and 0.4% for BR5, BR5CuL, and BR5CuH steels, respectively. Species from this genus are strictly aerobic, and their eff ect on steel corrosion has not been reported. The proportion ofDesulfobacterincreased from 0.1% on BR5 steel to 2.7% on BR5CuL steel, and then decreased to 1.4% on BR5CuH steel. As a typical SRB genus,Desulfobacterincluded in class Deltaproteobacteria aff ects steel corrosion similar to genusDesulfovibrio.Epsilonproteobacterial genusArcobactertook a percentage of 0.4%, 2.2%, and 0.4% on diff erent steels with increased copper contents, and it shows an unusually wide range of habitats. Some species are aerotolerant, while some are obligate anaerobes.Meanwhile, its role in sulfur cycling is variable, and some species oxidize reduced sulfur compounds(Voordouw, 2011), while some produce sulf ide(Jyothsna et al., 2013). They are expected to result in a complex impact ofArcobacteron steel corrosion.Although the largest diff erence ofSphaerochaetaandLabrenziafrom class Spirochaetes and Alphaproteobacteria was 2.0% (BR5 steel: 0.1%,BR5CuL steel: 2.2%, BR5CuH steel: 0.6%) and 1.0%(BR5 steel: 0.2%, BR5CuL steel: 1.2%, BR5CuH steel: 0.7%) among diff erent steels, there has been no knowledge about their inf luence on steel corrosion.Zetaproteobacterial genusMariprofunduscan aff ect steel corrosion like other IOB, but the variation in proportion was small among diff erent steels (0.5%,0.9%, and 0.8% for BR5, BR5CuL, and BR5CuH steels). The proportion ofRhiobiumincluded in class Alphaproteobacteria was also close, and the values were 0.5%, 0.8%, and 0.7%, respectively.
The variation in the relative abundance of the top 20 most abundant genera among diff erent steels is presented more directly by diff erent colors in a hierarchically clustered heatmap (Fig.7). Genera are clustered according to their changes in relative abundance with copper contents, and the distance among diff erent genera in the dendrogram ref lected their similarity in those changes. Furthermore, the distance between BR5CuL and BR5CuH steels was smaller than the other two couples, and therefore the distribution of the 20 most abundant genera in these two steels is closer.
4 DISCUSSION
4.1 An adverse impact of copper introduction on corrosion rate
In natural seawater, corrosion of HSLA steels occurs via the electrochemical actions of anodic iron oxidation (reaction 1) and cathodic dissolved oxygen reduction (reaction 2). Fe2+and OH-are combined to form Fe(OH)2(reaction 3), and further oxidized to Fe(OH)3via reaction 4. As Fe(OH)3is not stable, it transforms to γ-FeOOH via dehydration (reaction 5).When γ-FeOOH contacts with steels directly or mediates with other conductors, it is reduced (reaction 6) to favor corrosion. BR5, BR5CuL, and BR5CuH steels corroded in a similar way, evidenced by the γ-FeOOH and Fe3O4components in corrosion products from XRD results (Fig.3):
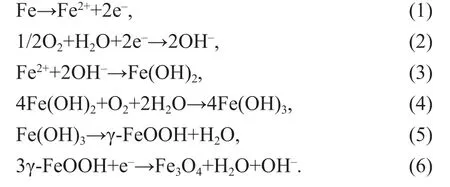
Copper introduction was detrimental to corrosion rate of mooring chain steels investigated in the present work, and the adverse impact was greater with 0.4%copper than that with 0.8% (Table 3). This conf licted with the report from Shi et al. (2018) that as-aged copper-bearing X80 pipeline steel gave better corrosion resistance against SRB than bare X80 steel,which might be closely associated with the variation in composition and microstructure of steels and characteristics of electrolytes. The adverse impact of copper alloying on long-term marine immersion corrosion of HSLA steels has been reported by Forgeson et al. (1960). Melchers (2004) proposed that the adverse impact of copper alloying was due to corrosion promotion at the anaerobic steady corrosion stage. Normally, it takes several years to get anaerobic corrosion developed completely, which is dependent on the average seawater temperature, and the present exposure period (three months) was too short to the arrival of that stage. Consequently, the detrimental role of copper alloying could not be attributed to its inf luence on anaerobic steady corrosion, and the mechanisms will be discussed below.
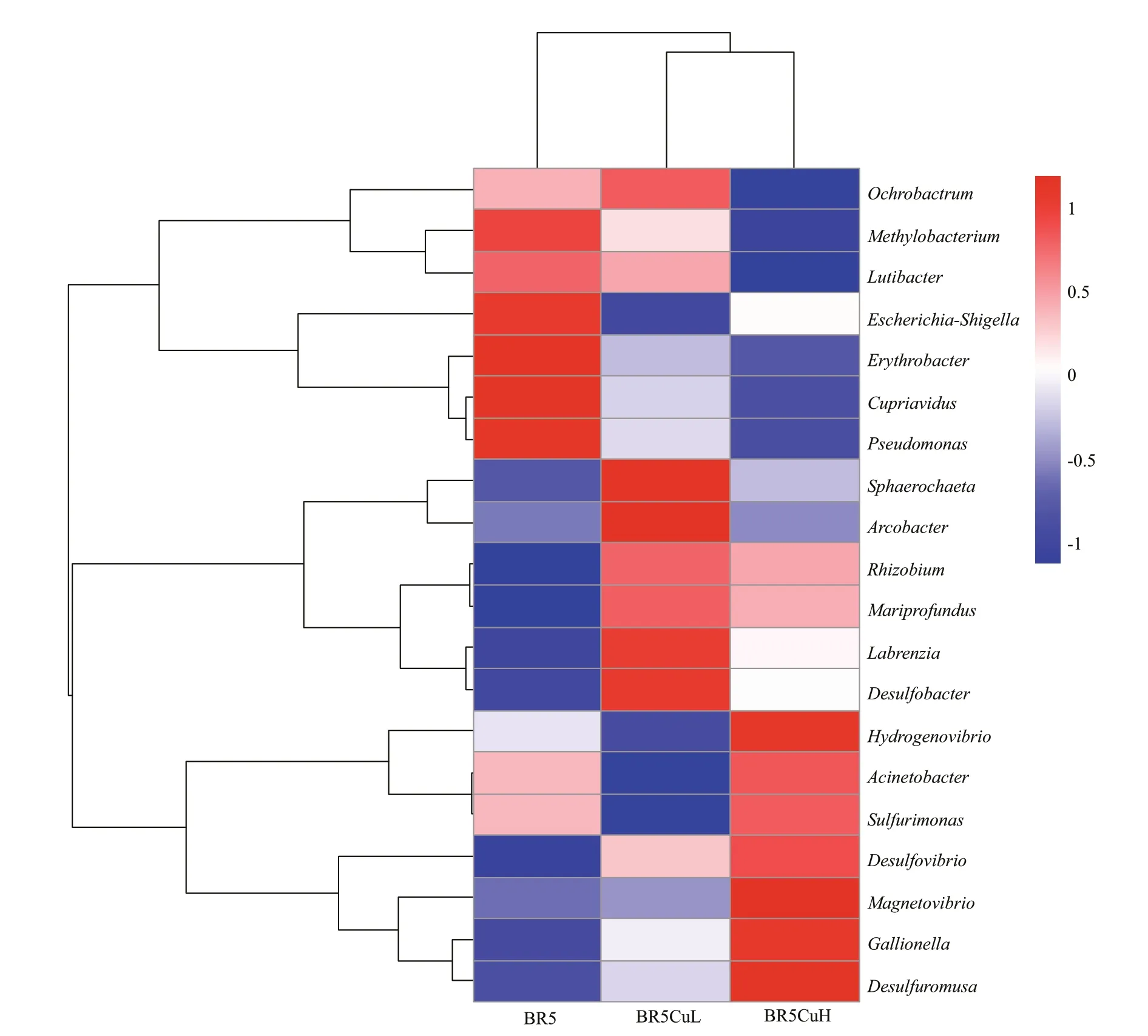
Fig.7 Double hierarchical dendrogram showing the 20 most abundant genera among microbial communities formed on diff erent steels
The dependence of corrosion rate on copper contents in the present work was diff erent from most cases that corrosion parameters increased or decreased linearly with the increased copper contents (Southwell and Alexander, 1970; Blekkenhorst et al., 1986).Unfortunately, these references available did not explain the causes of corrosion variation among steels with diff erent copper contents. Therefore, it is now diffi cult to discuss the diff erence between them and our results in a penetrating way, and we will just explain the variation among the present three steels.
4.2 Dependence of microbial community diversity,richness, and composition on copper contents
Unlike the case for most single strains that bacterial quantity was decreased by the introduction of copper into steels (Sun et al., 2016; Xu et al., 2018), BR5CuL and BR5CuH steels held a similar quantity to copperfree BR5 steel (Table 3). This diff erence could be ascribed to the stronger defense capacity against copper of natural complex consortia than that of pure strains, and the number loss in copper-sensitive species could be remedied by those with high resistance against copper like species from genusDesulfovibrio. The evolution of microbial community structure with copper introduction is discussed later.Diff erent from microbial quantity, microbial community diversity, and richness varied among diff erent steels (Table 4, Fig.4). BR5CuL steel provided biof ilms with the highest community diversity and richness, and BR5CuH gave the lowest.Copper is an essential trace element for organisms,and more than 30 types of copper-containing proteins are known today. However, excess copper is highly toxic, and this virucidal property has been exploited by humankind for centuries. Copper ions can be released from copper-rich phases in copper-bearing steels (Sun et al., 2016; Shi et al., 2018), and their amount was expected to be higher on the surface of BR5CuH steel than that on BR5CuL steel. When the concentration of copper ions is above the threshold,they become toxic to microbes, which was the case for BR5CuH steel in the present study. This decreased microbial community diversity and richness was in good accordance with extensive reports on microbes in aquatic environments and soils aff ected by copper ions with high concentrations (Smit et al., 1997; Wang et al., 2007), which derived from unavailability of certain copper-sensitive bacteria. The inhibition of copper towards microbe survival can be achieved by several mechanisms like replacing other metals in the metal-binding sites to interfere with the correct functioning of proteins (De la Iglesia et al., 2012),depolarizing and impairing of receptors or transporter molecules (Webster et al., 2001), and catalyzing the production of reactive oxygen species by a Fentonlike reaction (Dupont et al., 2011).
The higher microbial community diversity and richness on BR5CuL than that on BR5 steel might not be expected, because it was common in numerous reports that microbial community diversity and richness decreased with copper contents (Cantera et al., 2016; Taylor and Walker, 2016). However, Xie(2010) pointed that the inf luence of copper on microbial diversity and richness was not a simple linear relationship with its concentrations, and copper could be benef icial in a certain content range. In addition, this was proved by the work of Wang et al.(2008), who found that there was a signif icant increase in the number of PCR-denaturing gradient gel electrophoresis bands in soil withElsholtziasplendensincorporated with either 200 or 500 mg/kg copper,and the number of bands decreased sharply when copper concentration increased to 1 000 mg/kg. This increase in microbial diversity and richness was believed to be closely related to the stimulation of copper-adapted bacteria in response to the copper stress. According to the Venn diagram in Fig.2, there were 415 unique OTUs in biof ilms formed on BR5CuL steel, and some of these unique species are well known for their copper tolerance such as those from the generaDethiosulfatibacter,Desulfotomaculum, andDesulfotignum(data not shown).
Additional to microbial community diversity and richness, the relative abundance of common OTUs varied among diff erent steels (Figs.6 & 7). This microbial community structure alternation was universal in previous reports on the microbial response to the presence of copper, which was due to speciesdependent tolerance against copper (Zhao et al., 2014;Wang et al., 2015). It is important to mention that the response of individual genera or species may vary in diff erent reports, depending on copper speciations,copper contents, and the environments. In the present study, it seemed that generaPseudomonas,Cupriacidus,Lutibacter,Erythrobacter, andMethylobacteriumwere susceptible to copper stress evidenced by the decreased percentage with an increase in copper contents. AlthoughPseudomonashas been reported previously as one of the most copper-tolerant genera due to its capacity in the production of EPS having a high affi nity for copper ions (Li and Ramakrishna, 2011; Andreazza et al.,2012), it is not the case here. There might be two reasons for the continuous reduction in its relative abundance. Firstly, unlike the case of soil or nonmetal materials, mooring chain steels corroded with the consumption of oxygen under aerobic conditions,leading to an oxygen stress to the growth ofPseudomonas. Secondly, species with higher coppertolerance like those from genusDesulfovibriowere stimulated, which competed withPseudomonasfor nutrients, and their metabolites imposed great environmental pressure onPseudomonas. These two reasons also applied to the other 4 genera.
By contrast, generaDesulfovibrio,Desulfurumusa,Magnetovibrio, andGallionellapossessed higher copper resistance, and gave increased relative abundance with copper contents. The coppertolerance ofDesulfovibrioandDesulfurumusais closely related to their biogenic sulf ide, which binds with copper ions to form highly stable copper sulf ides with quite low solubility product constants (Utgikar et al., 2003). The formation of metal sulf ides dramatically reduces the bioavailablity of copper ions, and therefore copper toxicity is reduced.Meanwhile, EPS produced by species of these two genera are good copper-complexing ligands, which further decreases copper ion concentration and the toxicity (Flemming and Trevors, 1989). According to references available, the copper sensitivity of genusMagnetovibriois unclear. As a typical genus for SOB,its stimulation by copper introduction may be ascribed to the suffi cient sulf ide provided by the tremendous growth of SRB. Species from genusGallionellaare common in copper bioleaching industry due to its capacity in transferring Fe2+to Fe3+, which catalyzes the transformation of zero-valence copper to copper ions (Xiang et al., 2010). In turn, the transformation of copper provides new Fe2+forGallionella.Consequently, it was not surprising that copper introduction in mooring chain steel stimulated the growth ofGallionella.
Unlike the linear change of above nine genera with copper contents, generaSulfurimonas,Escherichia-Shigella,Hydrogenovibrio, andAcinetobacterpossessed the lowest relative abundance on BR5CuL steel. As typical genera for SOB, the percentage ofSulfurimonasandHydrogenovibriowas expected to improve due to the stimulation of SRB by copper introduction, which was in good agreement with their highest proportion on BR5CuH steel. The lowest relative abundance of these two genera on BR5CuL steel might be due to the promotion from SRB was smaller than the dispersion eff ect from the highest microbial community diversity/richness. Species from genusEscherichia-Shigellawere sensitive to copper, because the highest relative abundance was present on BR5. Similar toSulfurimonasandHydrogenovibrio, genusAcinetobacterexhibited the lowest and highest percentages on BR5CuL and BR5CuH steels, respectively. The highest relative abundance on BR5CuH steel was closely related to its well-known metal-tolerance via the expression of copper resistance proteins and the binging of EPS with metal ions (Yadav et al., 2013), and the lowest on BR5CuL steel could be ascribed to the predominant dispersion eff ect from the high community diversity/richness.
The other 7 genera gave the highest relative abundance on BR5CuL steel, demonstrating that they could tolerate copper and be stimulated at a low copper concentration, and the thresholds for copper toxicity ought to be lower than those ofDesulfovibrio,Magnetovibrio, andAcinetobacter, etc. Among them,genusOchrobactrumhas been reported to resist copper toxicity by several mechanisms, including binging copper ions on cell wall surface, generating EPS to chelate copper ions, bio-transporting copper from intramembrane to the outer membrane, and reducing copper with the aid of enzyme-mediated biotransformation (Ozdemir et al., 2003; Peng et al.,2019). The adaptation ofDesulfobacterto copper is similar to other SRB genera. The copper-tolerance mechanisms of the residual f ive genera are unclear due to quite limited references available.
4.3 Probable involvement of well-known corrosive bacteria and microbial community diversity/richness in corrosion acceleration
Bacteria involved in sulfur and iron cycling have long been viewed to play a signif icant role in steel corrosion, and the relative abundance of these special categories was analyzed. As described in the results part, there were typical genera for SRB (including sulfur-reducing bacteria), SOB, and IOB among the 20 most dominant genera. The sum of typical SRB generaDesulfovibrio,Desulfuromusa, andDesulfobacterwas 3.9%, 27.1%, and 35.6% on BR5, BR5CuL, and BR5CuH steels, respectively. Typical SOB also covered three genera, ieSulfurimonas,Hydrogenovibrio,andMagnetovibrio, and the total proportion was 12.0%for BR5 steel, 6.3% for BR5CuL steel, and 16.6% for BR5CuH steel. Besides, species from genusArcobactercould behave as SRB and SOB depending on the environments, and its proportion was 0.4%, 2.7%, and 1.4% for steels in the order of BR5, BR5CuL, and BR5CuH steels. IOB consisting of generaGallionellaandMariprofunduaoccupied 0.6%, 1.7%, and 2.6% of microbial communities on BR5, BR5CuL, and BR5CuH steels, respectively. It was obvious that SRB and IOB were stimulated by copper introduction, and SOB was restrained on BR5CuL steel, stimulated on BR5CuH steel. Combined with corrosion rate results, it could be found that BR5CuH steel with the lowest microbial community diversity/richness but the largest percentage in SRB, SOB, and IOB corroded more severely than BR5 steel, and consequently these typical well-known corrosive bacteria might be involved in the accelerated corrosion of BR5CuH steel. However,corrosion rate of BR5CuH steel was smaller than that of BR5CuL steel with a lower proportion of those corrosive bacteria but the highest microbial community diversity/richness. Therefore, only high relative abundance of well-known corrosive bacteria could not guarantee the high corrosion rate of BR5CuL steel, and complex microbial community structure ought to be considered in corrosion acceleration.
Extensive eff orts have been made to establish the relationship between corrosion rate and the amount or relative abundance of typical well-known corrosive bacteria, especially SRB, and there are some controversies. For example, Bonifay et al. (2017)compared microbial communities formed on pipeline systems with diff erent corrosion rate, and found that samples with high corrosion rate gave a rather high relative abundance of SRB, while Génin et al. (1994)reported that there was no signif icant diff erence in SRB quantity determined by the most-probable number technique at accelerated and normal low-water corrosion sites of steel sheet piles in a channel harbor.These conf licts may be closely related to the diff erence in characterization methods for microbial communities,exposure environments and time, and feature of substrates. Results from the present study seemed as a compromise for such conf licts, and a certain amount of well-known corrosive bacteria seemed to contribute to corrosion, but the corrosion rate did not rise linearly with their quantity. Furthermore, several reports have indicated that the metabolic status of SRB might be a relevant parameter to corrosion rate (Marty et al.,2014), and the metabolic activity of microbial communities will be investigated in our future work.
It is common that the corrosivity of natural microbial consortia with high diversity/richness is much stronger than that of single strains, which is associated with the synergy among diff erent species (Rao et al., 2000).Much attention has been paid to the analysis of microbial community components and their relationship with corrosion rate, and quite few reports dealt with the role of community diversity/richness. Although the diversity/richness depended on the composition of microbial communities, it could give an overall evaluation besides the well-known corrosive bacteria.In the present study, the microbial communities with the highest diversity/richness on BR5CuL steel held a large variety of species other than SRB, SOB, and IOB,and these species might contain highly corrosive bacteria that have not been recognized. With persistent eff orts in MIC research, more species are found to be highly corrosive by unique mechanisms, and a typical example was aMethanobacterium-like methanogen which captured electrons from iron via the direct electron transfer to accelerate corrosion greatly (Dinh et al., 2004). The involvement of community diversity/richness in corrosion helps cover the impact of potential corrosive bacteria. Furthermore, the high diversity/richness of microbial communities off ers a possibility to the complicated collaboration among diff erent species, which may also accelerate corrosion. Corrosion acceleration by the synergy of mixed species could be achieved by several modes like stimulation of anaerobes by oxygen consumption from aerobes (Wu et al., 2016), mutual stimulation of microbes metabolizing diff erent valences of the same element(Beech and Campbell, 2008), and promotion in electron transfer of some species by redox mediators from others (Zhang et al., 2015). Consequently, diversity/richness analysis ought to be involved together with structure in microbial communities related to corrosion.
5 CONCLUSION
Corrosion rate varied among three mooring chain steels with diff erent copper contents, increasing in the order of BR5 steel (without copper) < BR5CuH steel(0.8% copper) < BR5CuL steel (0.4% copper). The adverse impact of copper introduction on corrosion rate seemed to be closely related to the feature of microbial communities. Although copper introduction did not result in an obvious change in microbial quantity, it altered the diversity, richness, and structure of microbial communities due to species-dependence of tolerance against copper. The addition of copper stimulated the growth of some well-known corrosive bacteria like SRB and IOB, increased the community diversity/richness at a low content (0.4%), and reduced the diversity/richness sharply with the content of 0.8%. The highest corrosion rate of BR5CuL steel indicated that both well-known corrosive bacteria and microbial community diversity/richness might be involved in the corrosion acceleration. The present work shed some light on corrosion mechanisms of copper-bearing steels from the viewpoint of microbial communities, and there are still some defects like the lack of metabolomic analysis, which will be remedied in our future work.
6 DATA AVAILABILITY STATEMENT
The 16S rRNA gene sequences determined in this study have been submitted to the NCBI database with the accession number PRJNA493882.
 Journal of Oceanology and Limnology2020年2期
Journal of Oceanology and Limnology2020年2期
- Journal of Oceanology and Limnology的其它文章
- Contribution of surface wave-induced vertical mixing to heat content in global upper ocean*
- Upper ocean response to typhoon Kujira (2015) in the South China Sea by multiple means of observation*
- Inf luence of simulating deep-sea environmental factors on cathodic performance of seawater battery*
- Adsorption characteristics of chitooligosaccharides onto activated charcoal in aqueous solutions*
- Eff ects of hypoxia on survival, behavior, and metabolism of Zhikong scallop Chlamys farreri Jones et Preston 1904*
- Distinct inf luence of trimethylamine N-oxide and high hydrostatic pressure on community structure and culturable deep-sea bacteria*