
Eff ects of hypoxia on survival, behavior, and metabolism of Zhikong scallop Chlamys farreri Jones et Preston 1904*
2020-03-19 12:30LIQiaoZHANGFangWANGMinxiaoLIMengnaSUNSong
LI Qiao ZHANG Fang WANG Minxiao LI Mengna SUN Song
1 Key Laboratory of Marine Ecology and Environmental Sciences, Institute of Oceanology, Chinese Academy of Sciences,Qingdao 266071, China
2 University of Chinese Academy of Sciences, Beijing 100049, China
Abstract Hypoxia, a frequent occurring threat in coastal regions, often results in mass mortalities of marine organisms and brings a serious ecological problem. The commercially important Zhikong scallop Chlamys farreri is being under such a threat as the risks of eutrophication and hypoxia have risen in their culture areas. However, little information has been known concerning their tolerance to hypoxia and their strategy for survival. In the present study, a 20-day experiment was conducted to determine the eff ects of hypoxia on the survival, behavior, and metabolism of Zhikong scallop. With the LC 50 for dissolved oxygen (DO) being estimated as 1.8 mg/L, the survival of Zhikong scallop can be greatly challenged even under the moderate hypoxic condition of around 2.0 mg/L DO. The survival rate ranged from 69% to 59% when DO dropped from 3.0 to 2.0 mg/L, and it was further reduced to 47% at 1.5 mg/L DO. In hypoxic conditions, the scallops became signif icantly active, which may be explained as escape attempts to avoid hypoxic water. To save energy, Zhikong scallop would depress their respiration. However, when DO dropped from 3.0 to 2.0 mg/L,the oxygen consumption rate hardly changed. The upregulation of lactate dehydrogenase activity and the unrepressed phosphofructokinase activity, which often result in the unbalanced cellular homeostasis and energy budget, may account for the observed increase in the mortality rate of Zhikong scallops. In general,Zhikong scallop is sensitive to hypoxia events, though possible escape attempts, depressed respiration, and oxaloacetate-pathway may increase their survival chance.
Keyword: hypoxia; Zhikong scallop; survival; behavior; metabolism
1 INTRODUCTION
Zhikong scallopChlamysfarreri, a sessile and f ilter-feeding benthic animal, is widely distributed along the coast of northern China, and constitutes one of the major cultured bivalves due to its high nutritional value (Guo and Luo, 2006). In the past 30 years, both the culture area and density have soared,resulting in the deterioration of the environment.Mass mortalities were recorded several times during summer since 1996 (Zhang and Yang, 1999; Xiao et al., 2005; Wang et al., 2018). In an eff ort to determine the possible reasons accounting for the increase in mortality rates of scallops, several hypotheses have been proposed. Among these, pathogens including parasitic organisms and viruses are considered crucial factors. Additionally, the rising sea temperature has been proven an impact on immunity of the scallops(Chen et al., 2007b; Tang et al., 2010; Getchell, 2006).In addition, lower heterozygosity caused by inbreeding, toxic dinof lagellate, and shortage of food are also considered to be potential factors contributing to the severe mortality (Yan et al., 2001; Yang et al.,2001; Xiao et al., 2005). However, as a growing stressor worldwide and often a consequence of highdensity stocking and water quality deterioration, the eff ects of hypoxia has not been extensively studied.
Hypoxia is lethal to many marine organisms as mass mortalities often occur under severe hypoxia(Vaquer-Sunyer and Duarte, 2008). Hypoxic zones have signif icantly increased in the last few decades with the development of eutrophication and rising temperature (Diaz and Rosenberg, 2008; Howarth et al., 2011). In the culture area, the situation may become worse as the cultured organisms can produce large amounts of organic deposits, which may consume lots of oxygen in decomposition process. It is reported that in the Jiaozhou Bay, the culture of Zhikong scallops and bay scallops (Argopecten irradians) resulted in a decrease in dissolved oxygen(DO) levels to around 2.0 mg/L. In Sanggou Bay,another important culture area for Zhikong scallops,the risk of eutrophication and hypoxia have risen sharply due to aquaculture activities (Yang et al., 2018).
To survive hypoxia, metabolism is often depressed to save energy, as aerobic metabolism is limited when oxygen is insuffi cient. To save energy, the intensity of both physical and physiological activities, such as swimming, breathing, digestion, and heartbeat, are usually reduced to basal levels (Seibel, 2011;Breitburg et al., 2018). Furthermore, glucose, the main energy source for cells, can be broken down in multiple ways under anaerobic conditions: glucosesuccinate pathway, which features high effi ciency but low rates; lactate and opine pathways, which have low effi ciency but high rates. The former pathway is often considered the main strategy for hypoxiatolerant species due to the avoidance of acid metabolites, such as lactate, which may disrupt cell homeostasis (Larade et al., 2002). In addition, to control glycolytic f lux under hypoxia, covalent modif ication of key glycolytic enzymes often takes place. For example, the downregulation of lactate dehydrogenase (LDH) and pyruvate kinase (PK)activities inLittorinalittoreasignif icantly reduced its metabolic rate under hypoxia (Smolinski, 2016).
Generally, scallops are less tolerant to hypoxia than other bivalves. For example, the bay scallopArgopecten irradianscould only survive 8-24 h under the DO of 2-4 mg/L, although the DO is still suitable for most clams and oysters (Leverone, 1995). In addition to posing a great threat to survival, hypoxia is found to impart an inhibitory eff ect on the feeding, breathing,movement, and growth of bivalves. For example, the shell length and tissue dry weight of juvenile Green-Lipped MusselPernaviridissignif icantly decreased under hypoxic conditions, and the early developmental traits of the musselMytiluseduliswere adversely aff ected by reduced oxygen (Wang et al., 2011; Kong et al., 2019). Besides, it is reported that hypoxia often lead to the increasing hemocyte mortality and reactive oxygen species production in bivalves, which may leave them vulnerable to diseases and other threats (Sui et al., 2016). For biochemical responses, the glucosesuccinate pathway that is popular in many bivalves, has been conf irmed in the bay scallop,Argopectenirradians concentricus, based on the accumulation of succinate(Ivanina et al., 2016; Aguirre-Velarde et al., 2018).
For Zhikong scallop, mortality under moderate hypoxia has been reported, and several researchers have suggested that hypoxia may depress the immune responses of Zhikong scallop, increasing their sensitivity to pathogens (Chen et al., 2007a & c).However, none of these studies has investigated LC50levels to assess their tolerance to hypoxia precisely,and the response of key enzyme activities involved in respiration metabolism remains unclear, whether for Zhikong scallops or most marine benthos. Besides,most studies on the behavioral responses of bivalves to hypoxia have focused on the slow-moving or infaunal species. For example, theMacomabalthicawould reduce their burial depth to get more oxygen,and the anti-predatory responses ofPernaviridisandMytiluscoruscuswere often impaired by hypoxia(Long et al., 2008; Wang et al., 2013; Sui et al., 2016).In contrast, the behavioral responses of the free-living bivalves, including scallops, have not been studied extensively. Thus, to answer the questions that whether Zhikong scallop could survive a growing hypoxic threat and what kind of strategy it will adopt,a long-term experiment was conducted to determine the eff ects of hypoxia on the survival, behavior,physiological, and biochemical responses of Zhikong scallops. Determining the LC50for DO may improve our understanding of the consequences of highdensity culture and water deterioration and provide scientif ic advice for the scallop farming industry.
2 MATERIAL AND METHOD
2.1 Animals and laboratorial hypoxia

Fig.1 Schematic diagram of the experimental design
Zhikong scallop (mean shell height: 5.51±0.12 cm,mean weight: 23.86±2.79 g) were purchased from Mingsheng Farm, Qingdao, China on June 22, 2018.All of the scallops were transported to the laboratory within 2 h. During their transport, air was pumped continuously into the temporary containers f illed with seawater to avoid any possible hypoxia-acclimation.In the laboratory, all of the scallops were brushed carefully to remove attached debris on shells and were placed in several 250-L-aquaria (80-90 scallops in one aquarium). In the holding period,Chlorellaspp.(9.13±2.48 mg/L) were provided as food once a day.Water temperature was controlled at 18°C (average temperature of the bottom water in most of their habitats during summer). DO levels were monitored in real time using an OX11875 meter manufactured by Loligo Systems (Viborg, Denmark). Air stones were placed at the bottom of the aquaria, and high-pressure bottle was opened to pump compressed air into the aquaria when DO dropped below 7.0 mg/L. Ammonia nitrogen, hydrogen sulf ide, and pH were monitored daily using a multifunctional water quality analyzer HNT6S (Honart, Beijing, China), and one-third of the seawater was replaced every two days to ensure water quality. All of the aquaria were covered with a 2-mmthick black canvas to avoid light stimulation.
A circulating water system with DO- and temperature-controlled functions was designed for the experiments. Each DO concentration corresponded to a water supply tank and four replicate experimental tanks. The 800-L supply tank, which was equipped with a DO sensor, a temperature sensor, an ultraviolet lamp, a chiller, a heater, and air stones, served as the chamber to adjust DO and temperature of the sea water. The animals were housed in four 90-L experimental tanks, connected with the water supply tank. Between the water supply tank and the experimental tanks, a purif ier was installed to f ilter the impurities and metabolic wastes. The water was continuously pumped from water supply tank to the experimental tanks, and then f lowed back through the purif ier. Solenoid valves, which were controlled by rely and a computer, automatically opened and closed according to the actual value and set value of the DO:if the actual value is higher than the setting value,then nitrogen will be pumped in; if the actual value is lower than the setting value, then compressed air will be pumped in. The time it took for the DO sensors to reach the exact value and for the oxygen to diff use evenly in the tanks have been considered when designing the control program. The accuracy and stability of the DO in experimental tanks have been verif ied. The DO meter (OX11875) and sensors(OX11250) mentioned above were purchased from Loligo Systems (Viborg, Denmark). The purif iers were provided by Dalian Huixin Titanium Equipment Development Co., Ltd. (Dalian, China).
2.2 Survival and behavior observation
In the 20-day hypoxia tolerance experiment, DO concentrations were set as follows: 1.5, 2.0, 2.5, and 3.0 mg/L for the treatment groups, and 7.0 mg/L for the control group. In each group, four replicate experimental tanks were set. Among these, three tanks were used for survival, physiological, and biochemical studies, whereas the fourth tank was used for behavioral observation (Fig.1). The temperature was set to 18°C;salinity was controlled at 30.0±1.0; the f low rate of each experimental tank was set as 200 L/h. All of the experimental tanks were covered with 2-mm-thick black canvas to avoid stimulus from the sun, except for the tanks used for behavior observation.
Before the scallops were transported from the holding aquaria to experimental tanks, they were carefully examined and only the ones with intact shells, tentacles, and outer mantle were selected for the experiments. The selected scallops were then assigned randomly to the experimental tanks with 32 or 12 (behavior observation) in each. In the f irst 72 h,the DO levels in all of the experimental tanks were controlled at 7.0 mg/L to allow the scallops to adapt to the new environment, and any scallops that died or abnormally behaved (i.e., shells were tightly closed or tentacles were not stretched) during this period were removed. The DO levels in all of the tanks were then slowly and evenly reduced to the target value within 48 h and maintained for 20 days (from July 8 to 27).Animals were fed once a day withChlorellaspp.(6.37±2.87 mg/L) during the entire experiment. To maintain the quality of the seawater, one-third was replaced every other day. The DO levels in all of the experimental tanks were monitored daily using the Winkler method, and ammonia nitrogen, hydrogen sulf ide, and pH levels were analyzed every two days using the water quality analyzer earlier described.
In the pre-experiment, it is found that when a scallop is about to die under hypoxia, its shells will open and the outer mantle and tentacles will contract.Additionally, the gills often curl at some degree. Thus,during mortality check, the mantle and gill of the scallop that showed the symptoms described above would be touched using a glass rod for three times,and if the scallop did not respond, it would be considered dead. The dead scallops would be removed from the experimental tanks immediately to prevent it from destroying the water quality. Mortality was monitored every 12 h in all of the tanks during the acclimation period and the whole experiment.
For behavioral observations, f ive cameras were set to record the scallops in these specif ied tanks in real time. In addition, to better estimate the distance the scallops move, a vertical ruler was attached to the surface of the tank, and a cross-shaped ruler was installed at the bottom of the tank. Video camera master, a video recording software produced by ZG Technology Co., Ltd. (Tianjin, China) was adopted for video recording. The video f iles were then exported and analyzed to study the behavioral responses of the scallops. For each scallop, the horizontal and vertical moving distance were calculated, and the frequency of shell opening and closing was counted.
2.3 Physiological and biochemical measurements
To measure the oxygen consumption rate (OCR)continuously, a 30-L tightly closed respiratory chamber was designed (Fig.2). The water inlet was at the bottom of the cylindrical chamber, and the water outlet was at the top. A valve was connected to the water inlet to control the water f low. A DO sensor(OX11250) was placed in the middle of the chamber,and two stirrers were installed to ensure the oxygen to diff use evenly. Before the scallops were deployed in the chamber, their volume was calculated using water displacement method. During measurements, the sea water from water supply tank was intermittently injected into the chamber, and the DO at the end of each injection and the beginning of next injection were recorded. In addition, a blank chamber carrying no scallop was set at the same time to measure the oxygen consumption by other microorganisms. The OCR was calculated as follows:

Fig.2 Structure of the respiratory chamber

whereD1is the DO of the treatment chamber at the end of one injection, andD2is the DO of the treatment chamber at the beginning of next injection.D3andD4are the DO of the corresponding blank chamber at the same time whenD1andD2were measured.VCandVSrefer to the volume of the chamber and the scallops respectively.Nis the number of scallops in the treatment chamber, andtis the time interval between the measurement ofD1(D3) andD2(D4).
Ten respiratory chambers (diameter: 40 cm; height:18 cm) were set with each DO concentration corresponding to a chamber carrying initially 28 scallops and a blank chamber. All of the scallops were checked, selected, and handled the same way as described above. Each measurement lasted for about 10 min. The time interval between two measurements was 2 h during the period of DO-decreasing process and the f irst day the 20-day tolerance experiment. In the remaining 19 days, it was extended to 12 h.
Heart rate (HR) and enzyme activities were analyzed at the end of the 20-day tolerance experiment. For HR measurement, eight scallops from each group were randomly designated. The sensors were attached to the shells with nail-free glue. Powerlab 8/35 produced by AD instrument (New South Wales, Australia) was used for the signal collection and analysis. The measurements started 2 h after the deployment of the sensors to avoid disturbance caused by the handling. The waveform f iles were then analyzed and only the segments with clear and stable waveform were adopted for HR counting. One heartbeat cycle was def ined as the time from one wave crest to the next wave crest or from one trough to the next trough.
For enzyme activity measurements, the activities of phosphofructokinase (PFK), LDH,phosphoenolpyruvate carboxylase (PEPC), fumaric reductase (FR), and octopine dehydrogenase (ODH)were studied in the adductor muscle of the scallops.PFK is the most crucial rate-limiting enzyme in glycolysis. The activities of PEPC and FR usually ref lect the pathway of phosphoenolpyruvate metabolism and the anaerobic metabolism. LDH and ODH are the common pyruvate reductase in bivalves.Eight random scallops were selected from each group,and the adductor muscle of each scallop was cut into several 3 mm×3 mm×3 mm sections. Each section was designated for the measurement of one enzyme and was temporarily stored in a centrifuge tube in the icebox. To avoid the potential bias caused by subjective consciousness, the experimenter responsible for the following analysis was blinded from the group information as the centrifuge tubes were re-numbered by another experimenter. For the measurements of PFK, LDH, and PEPC activities,commercial assay kits (A129, A020-2, and A130;Nanjing Jiancheng Bioengineering Institute, Nanjing,China) were used according to the manufacturer’s instructions. For FR activity analysis, the method described in the Inhibition of Albendazole and Oxfendazole on the Activity of Fumaric Reductase inCysticercuscellulosae(Gao et al., 2002) was adopted,whereas for ODH, the method in Further Studies on the Phylogenetic Distribution of Pyruvate Oxidoreductase Activities (Livingstone et al., 1990) was employed.
2.4 Post eff ect analysis
Since hypoxic events may occur several times during the summer, frequent deoxygenation and oxygenation may bring extra oxidative damage to the scallops. To determine the survival rates of Zhikong scallops in a second hypoxic event, the DO of treatment tanks designated for survival observation were recovered to normoxic condition (7.0 mg/L)after physiological and biochemical sampling, and maintained for seven days. Then, the DO of all tanks,including the control ones in the previous tolerance experiment, were reduced to 1.5 mg/L evenly in 48 h and maintained for another 10 days. Other environmental settings and daily operations, including temperature, salinity, f low rates, sunshade canvas,and water changing frequency were the same as before. Mortality was monitored every 12 h.

Fig.3 Changes in survival rates of Zhikong scallop in the 20-day hypoxia tolerance experiment
2.5 Statistical analyses
Statistical analyses were conducted in survival rates, moving distance, frequency of shell opening and closing, HR, and enzyme activities (Supplementary Tables S1-S2). The data were f irst tested for normality and homogeneity of variances. If the data successfully pass the two tests, then one-way ANOVA was adopted,and if the data failed to pass any of the above tests,then Kruskal-Wallis H-test would be adopted to ensure the reliability of the conclusion, though the sensitivity of Kruskal-Wallis H-test is lower than ANOVA. Survival rates were analyzed using the latter method, as the Kruskal-Wallis H-test is reliable when the number of replicates are not large enough. Tukey test was used for multiple comparisons corresponding to one-way ANOVA, whereas the Mann-Whitney U-test was used when Kruskal-Wallis H-test was adopted. Statistical signif icance of the results was determined when aP-value of <0.05 was obtained.The 20-day LC50was estimated using probit analysis(Finney, 2009). All of the above analyses were conducted using SPSS 16.0 (IBM, Armonk, USA).
3 RESULT
3.1 Survival and behavioral response

Fig.4 Behavioral responses of Zhikong scallop to hypoxia
Hypoxia posed a great threat to the survival of Zhikong scallops, as the survival rate was reduced with the decreasing DO (Fig.3). The scallops exposed to 1.5 mg/L DO was apparently the most disrupted in this study, as the observed survival rate was <50%and was signif icantly lower than the other treatment groups. The survival rates of the treatment groups exposed to 2.0, 2.5, and 3.0 mg/L DO are relatively close, ranging from 59% to 69%. Mortality occurred nearly at the beginning of the 20-day tolerance experiment. However, after about eight days, the survival rates of all of the groups came to a relatively stable stage, which lasted about four days. Then, the survival rates continued to drop until the end of the experiment. The scallops of the control group were all alive when the experiment was over, suggesting that there was no other environmental threat but hypoxia in this variable-controlled experiment. The LT50at 1.5 mg/L DO was 432 h, and the 20-day LC50for DO was calculated to be 1.8 mg/L using probit analysis.

Fig.5 Changes in oxygen consumption rate of Zhikong scallop
Figure 4 shows that hypoxia has a signif icant eff ect on the horizontal and vertical movements of the Zhikong scallops. Except for the f irst 96 h, the scallops exposed to 1.5, 2.0 and 2.5 mg/L DO were more active in terms of horizontal movements. The treatment group of 3.0 mg/L DO and the control group did not diff er at most times. For vertical movements, the scallops of the treatment groups were quite active as the average moving distance often exceeded 160 cm per individual. The frequency of shell opening and closing did not diff er much across diff erent groups except that the control group showed a signif icant high value in the f irst 48 h.
3.2 Physiological responses
The OCR reduced immediately when the DO started to drop, and maintained relatively stable values after the target DO were reached. The lower the DO, the lower the OCR, though the diff erences between the treatment groups exposed to 2.0, 2.5, and 3.0 mg/L DO were small (Fig.5). During the 48-h oxygen-decreasing period, the OCR did not decrease.A phenomenon that the OCR initially decreased, then rose and then decreased again was observed among all treatment groups. In the following period of the 20-day experiment, the OCR of all of the groups only slightly f luctuated.

Fig.6 Heart rates of Zhikong scallop at the end of the 20-day experiment, under diff erent DO concentrations
The eff ect of hypoxia on heart rate was signif icant as the heart rate of the scallops exposed to 1.5 mg/L DO was depressed to around 20 beats per minute.However, there was no signif icant diff erence between the treatment groups exposed to 2.0, 2.5, and 3.0 mg/L DO and the control group (Fig.6).
3.3 Biochemical response
The eff ects of hypoxia on the activities of key enzymes involved in respiratory metabolism were signif icant (Fig.7). FR activity in adductor muscle of Zhikong scallops responded drastically to decreasing oxygen levels as it peaked under the DO of 3.0 mg/L and gradually dropped with further decrease in DO.The FR activity under 1.5 mg/L DO was roughly equal to that under normoxic condition. The changes in LDH and PEPC activities showed a similar trend as the lower level of DO resulted in an increase in enzyme activities. Notably, the LDH activity of the control group showed almost no activity. The PFK activity decreased when DO dropped below 2.5 mg/L(P=0.033). ODH activity did not respond to hypoxia as no signif icant diff erence was detected between the treatment groups and the control group.
3.4 Post eff ect analysis
After the DO in all of the treatment groups were reset to normoxic level, mortality rates increases in the groups exposed to the reduced DO in the previous experiment (Fig.8). At the end of the second hypoxia application, the survival rates of the groups once exposed to the reduced DO were 34%, 56%, 56%, and 55%, lower than that of the group once exposed to normoxia (71%). Besides, they were also lower than the survival rate on day 10 of the treatment group once exposed to 1.5 mg/L DO (67%). The treatment group once exposed to 1.5 mg/L DO had a minimum survival rate during the oxygen-decreasing period and the entire time of the second hypoxia.
4 DISCUSSION
4.1 Survival challenge
Hypoxia, a growing threat caused by eutrophication or high-density mariculture, signif icantly reduced the survival chance of Zhikong scallops, as shown in Fig.3. The 20-day LC50for DO was estimated to be 1.8 mg/L, indicating its tolerance to hypoxia was weaker than most other bivalves. For example, the Manila clamRuditapesphilippinarumis tolerant to 2 mg/L-hypoxia without mortality, whereas the Asian oysterCrassostreaariakensisand the Eastern oysterCrassostreavirginicacan survive more than 10 days under 0.5 mg/L DO (Lombardi et al., 2013; Zhang et al., 2017). Thus, to reduce economic losses, close DO monitoring was necessary and appropriate measures could be adopted to prevent the culture area from deadly hypoxia events.
An important phenomenon to notice is that the survival rate did not change linearly with DO. Figure 3 shows that the survival rate lines corresponding to the groups exposed to 2.0, 2.5, and 3.0 mg/L DO came pretty close, while the line corresponding to the group exposed to 1.5 mg/L DO was far apart. This may implicate there is an oxygen threshold between 1.5 and 2.0 mg/L for Zhikong scallop. It can be explained that the physical stress would break the limit of metabolic regulation when DO dropped below that threshold. In the study of another bivalve,Macomabalthica, the similar phenomenon was observed as mortality rates sharply increased when DO dropped below 2.5 mg/L (Long et al., 2008).
Another phenomenon worth paying attention to is that the survival rates did not always drop with time.This may indicate that after a certain time of hypoxia exposure, Zhikong scallops could adapt to the low DO concentrations to some degree. Furthermore, the time when mortality would come to a halt may represent the respond speed of adaptive mechanism.This phenomenon is quite meaningful, as it would protect Zhikong scallop population from complete destruction if DO concentration could rise after about 10 days. For another bivalve,Donaxserra, the same trend was also observed (Laudien et al., 2002). However,since the temporary stop of mortality would come too late,it is meaningless forDonaxserrapopulation to recover.


Fig.7 Biochemical responses to hypoxia measured in adductor muscle of Zhikong scallop
In the summer, some coastal and estuarine hypoxia often occur more than once as the suitable hydrographical condition for water stratif ication may form and rupture several times within months. Thus,the survival of marine organisms may be challenged repeatedly. As an organism that is able to live in the intertidal zone f ixing on rocks, Zhikong scallops are considered to have strong tolerance against the repeated deoxygenation and re-oxygenation processes like other intertidal benthos (Ivanina et al., 2016).However, in the present study, the scallops exposed to hypoxia for a second time have a survival rate 15% to 35% lower than those exposed to hypoxia for the f irst time. This result may again change people’s understanding about the hypoxia tolerance of those intertidal organisms. The scallop culturists should also monitor water quality and take measures to prevent f luctuations in oxygen levels.
4.2 Strategy to survive hypoxia
To increase survival chances under hypoxia,marine benthos would respond in terms of their behavior. Fish with strong swimming ability often initially try to escape hypoxic zones, and these attempts have been observed in some shrimps, such as the penaeid shrimpMetapenaeusensis(Wu et al.,2002; Domenici et al., 2007). Scallops, with highly developed adductor muscle, are capable of swimming a short distance by rapidly opening and closing their shells.
For horizontal movements, the scallops were particularly active during the f irst 48 h, which may be attributed to adaptation eff orts. In the following 18 days, the scallops exposed to hypoxia were generally more active than the scallops of the control group, which may be explained as the attempts to escape the challenging environments. However,although the diff erences were signif icant in statistics,the absolute values of the diff erence only ranged from about 40 to 50 cm, and it seemed useless for scallops to escape hypoxia in the wild. Here, we propose two assumptions: f irst, Zhikong scallop did attempt to escape hypoxia. However, the small experimental tanks limit their activity; another possible situation is that the increased horizontal movements were not due to the escape attempts, but due to the anxious response under hypoxia-stress. Thus, to further answer this question, it may be helpful to conduct further investigations in the wild or to conduct the experiments with huge tanks in the future.
Vertical movements in the control group were barely observed as most scallops were f ixed to the bottom of the tanks and only make minor horizontal movements around the f ixation points. The scallops exposed to hypoxia showed signif icantly high activity,with some individuals moving more than 240 cm.This fact seemed to support the f irst assumption described above, as the height of the experimental tank was longer than its diameter. From this perspective, Zhikong scallops may have a chance to avoid a hypoxic zone if the hypoxia is at its early stage or not spread to a large area. The frequency of shell opening and closing provides little implication.

Fig.8 Post eff ects of hypoxia
However, if it is impossible to escape from hypoxic water, metabolic regulation is crucial. As the effi ciency of anaerobic metabolism is much lower than that of aerobic metabolism, it is crucial to save energy when oxygen is insuffi cient. Oxygen consumption rate and heart rate, two important indicators of metabolic rate,are often depressed under hypoxia (Grieshaber et al.,1993; Gobler and Baumann, 2016). In 2014, Artigaud et al. (2014) studied the respiratory responses of the great scallop,Pectenmaximus. Compared to the Zhikong scallop, the great scallops seemed to have stronger ability in respiratory regulation as the OCR barely changed when DO dropped from 100% to 40%saturation. As shown in Fig.5, the OCR of Zhikong scallop decreased by more than a half when DO dropped from 7.0 to 3.0 mg/L. However, when DO further dropped from 3.0 to 2.0 mg/L, the OCR showed little response. This new strategy has rarely been reported. An advantage of this strategy could be that it is helpful for energy saving and storage, as feeding and digestion activities are often depressed under hypoxic conditions (Foss et al., 2002; Ripley and Foran, 2007). However, if an abnormal as well as depressed metabolism last for too long, then it may cause potential damage to the scallops.
In Fig.5, it is easy to notice another interesting phenomenon that during the OCR-decreasing process,there was a little peak when DO dropped to about 3.0 mg/L. The phenomenon that OCR increases with decreasing DO was once reported in some f ish. For example, the rainbow troutSalmogairdneriwas able to double its OCR under hypoxia. However, according to the conclusions, the utilization rate of oxygen reduced; the increased consumption of oxygen was not f lowing into the metabolism of the body. Instead,it was the result of added eff orts in breathing (Hughes and Sunders, 1970). For Zhikong scallops, the same thing could happen when they decide to regulate their respiration and prevent a further metabolic decline.To answer this question clearly, further investigations are warranted.
The cardiac response to hypoxia may diff er across diff erent species. In some shrimps,Nephrops norvegicusandMetapenaeusensisfor example, the HR were reduced with the decreasing DO (Hagerman and Uglow, 1985; Wu et al., 2002). However, in the scallopPectenmaximus, a gradually decreasing DO would result in a slight regulatory upswing of HR,and in the European sea bass,Dicentrarchuslabrax, a strong hypoxia-tolerance was often correlated with higher rates of cardiac contraction and relaxation(Brand and Roberts, 1973; Joyce et al., 2016). Figure 6 shows that there was no signif icant decrease in HR when DO dropped to 2.0 mg/L. Even at the most severe hypoxic group (1.5 mg/L DO), HR was only slightly reduced, suggesting the lack of cardiac regulatory ability of Zhikong scallops. An increasing HR could pump more blood as well as more oxygen per unit time. However, the increase in heart activity itself will cost more energy, thus requiring more oxygen. Thus, the result we see here was part of a larger energy balance scheme.
As the major energy-generating reaction under hypoxia, glucose metabolism often has diversif ied pathways. At the beginning of hypoxia, glycolysis is usually coupled with the transamination of aspartate.During the process, pyruvate converts into succinate,leaving oxaloacetate, malate, and fumarate as intermediates, partially reversing the tricarboxylic acid cycle. With the depletion of aspartate,oxaloacetate will be produced directly from phosphoenolpyruvate instead of pyruvate. (Larade and Storey, 2002). Figure 7 shows an increase in FR and PEPC under hypoxia, which may prove the existence of the pathway described above. However,after the FR activity peaked under 3.0 mg/L DO, it began to decrease with DO, which may indicate that the glucose-succinate pathway was not selected as the main strategy when oxygen is seriously insuffi cient.
Additionally, it has been reported that pyruvate often reacts with arginine to form octopine, instead of lactate in invertebrates (Hardewing et al., 1993). This metabolic pathway is considered crucial as it can avoid the accumulation of acids and maintain cellular homeostasis. However, according to the present study,the ODH (the enzyme that catalyzed the production of octopine) activity did not change when DO dropped from 7.0 to 1.5 mg/L. Conversely, the LDH was greatly activated with decreasing DO, which may result in the excessive accumulation of lactate. Thus,the extremely high LDH activity may give us possible clues relating to the death of Zhikong scallops under hypoxia. Besides, the changes of PEPC activity was consistent with LDH activity, given that H+is one of the activators of PEPC. Another important clue about the death of Zhikong scallop was that the PFK activity barely changed under hypoxia as it is often depressed to keep metabolism running at a low rate when oxygen is limited (Larade and Storey, 2002).
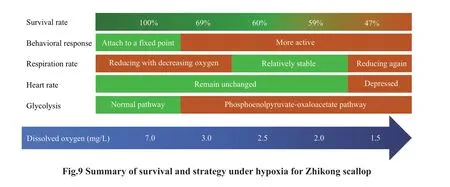
To better summarize the tolerance and response of Zhikong scallop under hypoxia, a summary f igure(Fig.9) is presented below.
5 CONCLUSION
To investigate the eff ects of hypoxia on survival,behavior, physiological, and biochemical response of the commercially important Zhikong scallop, this 20-day experiment was conducted, and the implications of the results follow: (1) the survival of Zhikong scallop would be greatly threatened even in a moderate hypoxia, and a DO concentration lower than 2.0 mg/L could cause the population to collapse if the hypoxia lasts more than 10 days; (2) if the hypoxic event was within a limited water column, then the Zhikong scallops have a chance to escape by swimming; (3)the respiration of Zhikong scallops would be depressed to reduce metabolic rate when DO began to drop, but stay relatively stable when DO further drop from 3.0 to 2.0 mg/L; (4) the signif icant high LDH activity and unrepressed PFK activity may be responsible for the f inal death of Zhikong scallops;(5) Zhikong scallops may become less tolerant to a second hypoxia event.
6 DATA AVAILABILITY STATEMENT
The datasets generated and/or analyzed during the current study are available from the corresponding author on reasonable request.
7 ACKNOWLEDGMENT
We thank DAI Li for providing the unicellular algae, and ZHOU Li for providing the instruments for enzyme-activity analysis. We are grateful for the assistance of YU Nan who helped us greatly in materials preparation.
 Journal of Oceanology and Limnology2020年2期
Journal of Oceanology and Limnology2020年2期
- Journal of Oceanology and Limnology的其它文章
- Contribution of surface wave-induced vertical mixing to heat content in global upper ocean*
- Upper ocean response to typhoon Kujira (2015) in the South China Sea by multiple means of observation*
- Inf luence of simulating deep-sea environmental factors on cathodic performance of seawater battery*
- Adsorption characteristics of chitooligosaccharides onto activated charcoal in aqueous solutions*
- Distinct inf luence of trimethylamine N-oxide and high hydrostatic pressure on community structure and culturable deep-sea bacteria*
- Microbial communities present on mooring chain steels with diff erent copper contents and corrosion rates*