
脂肪酶的生物学改造研究进展
2020-03-12 01:06王玉娟
生物学杂志 2020年1期
叶 纯,王玉娟
(1. 中国科学院 强磁场科学中心,合肥 230031; 2. 中国科学技术大学,合肥 230026)
脂肪酶作为工业生产中重要的生物催化剂[1],具有催化特异性高、反应条件温和、副产物少及环境友好等特点,但是脂肪酶制剂价格昂贵、酶与底物难分离、多余游离脂肪酶难回收等问题制约了其工业应用。因此,对天然脂肪酶进行针对性改造是当前本领域研究热点。本文将在简介脂肪酶的分子结构、催化机理等基本信息的基础上,综述利用定向进化等生物技术对其进行改造的研究现状,展望脂肪酶生物学改造的发展趋势,助益同行学者开展相关研究。
1 脂肪酶的概述
脂肪酶(Lipase,E.C. 3.1.1.3)又称酰基甘油水解酶,广泛来源于多种动植物及微生物。它能够催化水解、酯化、醇解等生化反应[2]。脂肪酶在催化反应中具位点选择性、立体选择性及化学选择性的特点,催化过程不需要辅因子,且具有高催化活性[3]。
1.1 脂肪酶结构
1990年,Brad等解析了米黑根毛霉脂肪酶(Rhizomucormieheilipase,RML)的晶体结构[4],到目前为止,已经有数百种脂肪酶的结构被解析出来。脂肪酶结合底物不同,或者所处溶剂环境不同,都可能呈现出不同的构象状态。不同物种的脂肪酶一级结构(氨基酸序列)相似度很低,且蛋白分子量也相差甚远,从16.25 ku(Rhizomucororyzaede的脂肪酶)[5]到116.6 ku(人体的激素敏感脂肪酶)[6],但它们的活性中心在二级和三级结构上相对保守,都具有相似的α/β水解酶折叠结构,是一个催化三联体结构(由丝氨酸、组氨酸、天冬氨酸组成)[2]。
1.2 脂肪酶的催化机理
脂肪酶的催化过程主要分为4个阶段,如图1所示:1)疏水性底物进入反应体系时,脂肪酶由α螺旋组成的盖子打开并暴露反应活性位点,底物进入活性中心,与脂肪酶结合。2)脂肪酶活性中心上的丝氨酸被活化,羟基氧原子对底物酯键上的羰基进行亲核攻击,羰基双键断裂,4种不同原子与该羰基碳原子键合,形成四面体中间复合物。3)四面体中间复合物发生扭转,活性中心的组氨酸将其从丝氨酸上获得的质子传递给底物酯键上的羟基,底物酯键断裂,生成的醇羟基部分脱离中间复合体,羰基部分与活性中心的丝氨酸形成新的酯键,酶与底物形成了一种酰基-酶共价中间复合体形式。4)脱酰基。活性中心的组氨酸失去质子以后,从底物分子水或醇中获取质子,底物分子水和醇被活化,攻击丝氨酸上的碳原子,丝氨酸与底物结合的酯键断裂,释放其羧基部分的产物(水解释放酸,醇解释放酯),同时脂肪酶完成了去酰基化过程[2,7]。
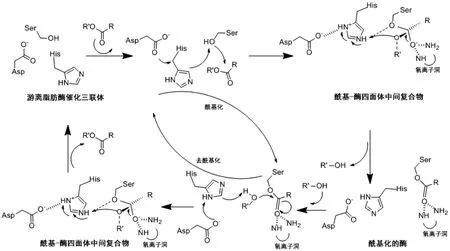
图1 脂肪酶催化机理[2]
2 脂肪酶的生物学改造
由于酶催化的底物分子专一性强,且受pH、温度等反应条件的限制,限制了其工业应用。因此,利用生物学技术改善酶的工业生产应用性能,成为近年来的研究热点。目前,计算机模拟、序列分析、基因突变与合成以及蛋白质工程等技术交叉融合,发展出定向进化、理性设计、从头设计和蛋白质表面修饰及固定化技术,使酶改造进入了主动时代,对脂肪酶催化特性的改造更具有针对性[7]。
2.1 定向进化
定向进化是在体外的实验环境中来模拟生物体自然进化的实验手段,通过随机突变氨基酸,构建突变重组文库,再进行定向筛选,以得到预期的目的蛋白,定向进化在不了解酶蛋白的结构及催化机制的条件下,也可以有目的地获得具有特定催化性能的突变体酶蛋白[8]。在定向进化中,构建突变重组文库的手段主要包括易错PCR[9]、DNA改组[10]、寡聚核苷酸重组(Assembly of designed oligonucleotides,ADO)[11]等,例如黄瑛[12]利用该技术对来源于短小芽孢杆菌的脂肪酶基因BpL进行定向进化,获得了BpL1-7和BpL2-1369两种突变菌株,其催化活力较野生型分别提高了2倍和6倍。苏宏飞等[13]利用DNA改组技术对脂肪酶Lip98进行定向改造,获得两个突变体F92A和I199F,其热稳定性与野生型相比分别提高了1.5和3.5倍。常用的高通量筛选方法主要有琼脂平板筛选、特异性酶产物筛选、表面展示技术等,例如Emmanuel等[14]利用脂肪酶催化水解反应,生成一种稳定性的荧光基团伞形酮的特性,观察产物荧光强度的变化来反映脂肪酶酶活,从而筛选出高酶活脂肪酶。定向进化方法高度依赖于完善的突变重组文库以及高效的高通量的定向筛选方法,存在着无效突变多、筛选困难及工作量大等问题。
2.2 理性设计
酶的理性设计是根据脂肪酶的已知结构信息及催化机理,对脂肪酶的结构与功能之间的关系进行分析,找出影响酶的稳定性、催化活性、折叠方式等的主要位点,然后对其进行针对性的修饰或替换,以改变脂肪酶的相应性状[15]。传统的理性设计方法主要基于实验以及序列分析,近年来,计算机辅助设计被广泛应用于脂肪酶的理性设计中。通过序列比对分析氨基酸序列的保守性,利用同源建模方法对未知蛋白进行模型构建,得到蛋白结构的理论模型[27]。同时根据蛋白质与配体之间的柔性对接以及刚性对接,发展出分子对接的计算分析方法,分析蛋白质与配体的结合自由能变化,以评价蛋白质的配体选择性。通过分子动力学模拟对蛋白质与配体结合体进行分析和预测,分析环境体系等因素对其结构及相互作用的影响,得到新型酶蛋白的相关催化活性、稳定性以及底物选择性等理论数据,从而选择最合适的突变体并指导实验,得到目标酶蛋白[17]。
2.3 半理性设计
由于随机突变的定向进化技术构建出的突变体文库数量巨大,筛选困难,而理性设计需要对脂肪酶结构、催化机理以及功能有深入的了解,限制了脂肪酶的改造。基于上述两种改造方法建立的半理性设计,即先用理性设计的思路,利用生物信息学手段对目标脂肪酶已有信息进行分析,将定向进化的突变位点锁定在有限几个氨基酸上,大大缩小了突变体文库的数量,且有效突变率大大增高[18]。例如,Reetz课题组基于脂肪酶有义突变主要集中在催化活性位点附近这一机理,提出了迭代饱和突变法(Iterative saturation mutagenesis,ISM)[19],先分析酶的结构、序列信息,选取活性中心关键位点,分别对这些位点进行饱和突变,从中筛选有义突变,对这些突变体再进行饱和突变,通过这样的多轮迭代饱和突变,找到最适突变体。
2.4 蛋白修饰
为了弥补氨基酸定点突变只能选择19种氨基酸对其进行替换的不足,蛋白质修饰方法被应用于酶蛋白改造中,使酶蛋白在目标位点上的改造更多样。脂肪酶蛋白的表面修饰主要通过对主链的剪切、连接以及对侧链氨基酸残基或者某个官能团进行化学修饰来实现。通过对酶蛋白进行表面修饰,可以增强其在不利催化环境中的稳定性及催化活力。如Abuchowski等的研究表明,聚乙二醇(PEG)可以通过改变脂肪酸的主链结构或侧链基团对其修饰,使脂肪酶在有机溶剂中更稳定,溶解性更强[20]。
2.5 固定化
在自然条件下,游离酶极易失活且回收困难,所以脂肪酶通常需要进行固定化操作才能应用于工业生产中,固定化的酶具有更高的环境耐受性及稳定性[21],利于回收再利用。目前工业生产上广泛应用的脂肪酶固定方法主要有4种:交联法、包埋法、吸附法以及共价法[22]。新型固定化技术,如纳米载体固定化利用纳米载体,包括纳米片、纳米纤维、纳米管等,对脂肪酶进行固定化(表1)[22]。定向固定化技术通过使酶蛋白活性位点远离固定化载体,使底物更容易与脂肪酶结合,提高了催化效率[7]。例如Godoy等[23]突变了来源于Geobacillusthermocatenulatus的脂肪酶BTL2,突变体φ-BTL2-S334C通过引入的半胱氨酸与固定载体上的醛基反应,将酶固定在二硫化物-醛支持物上(图2),使其在水解中具有几乎完全的选择性。
3 几种改造技术比较
通过对目前脂肪酶主要改造策略进行分析,总结了几种策略的主要技术特点。如表1所示,每种技术都有自己一定的局限性,多种技术相结合对脂肪酶进行改造成为未来研究的重要方向。
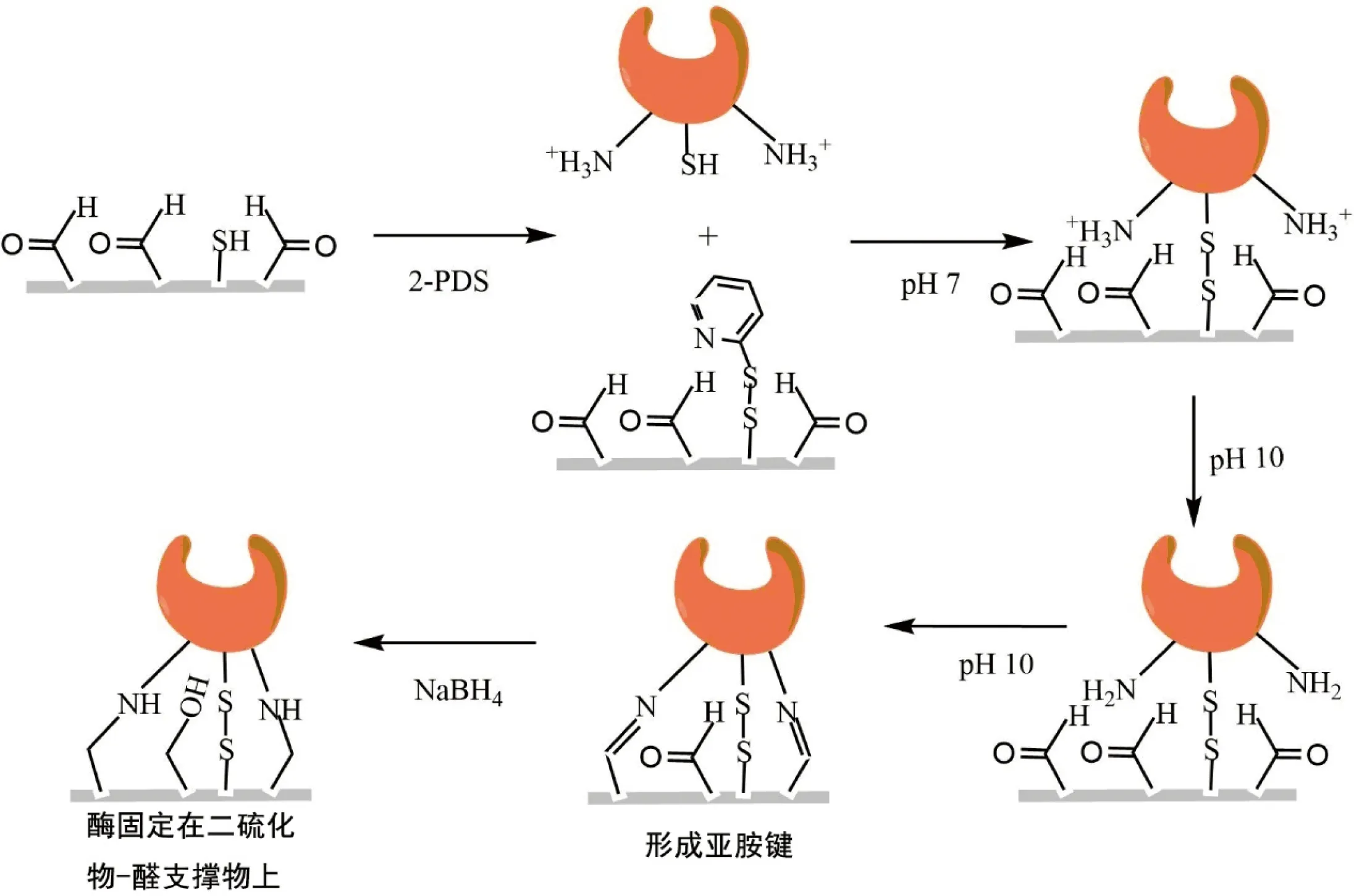
图2 φ-BTL2-S334C定向固定化流程[23]

表1 几种重要脂肪酶改造技术的比较
4 总结与展望
随着大量的研究,人们对脂肪酶的结构、功能和催化机理的了解越来越深入,为脂肪酶改造提供了理论支持。计算机硬件及相关软件越来越完善,为脂肪酶的改造提供了方向上的指导,提高了改造效率。本文总结了现有用于脂肪酶改造的生物技术,包括定向进化、理性设计、蛋白修饰、固定化等,并系统地分析了相关技术的优劣势。展望未来,现有多种技术相结合,优势互补以及各种新型改造技术的开发和运用将会极大地促进脂肪酶改造研究的发展。
猜你喜欢
中国农业科学(2022年12期)2022-06-28
中国畜牧杂志(2022年1期)2022-01-20
南昌大学学报(医学版)(2021年1期)2021-11-29
农业科技通讯(2021年1期)2021-03-06
中国洗涤用品工业(2017年2期)2017-04-16
上海农业学报(2017年3期)2017-04-10
天然产物研究与开发(2016年1期)2016-06-05
中国塑料(2016年7期)2016-04-16
天津医科大学学报(2015年2期)2015-12-22
中国粮油学报(2014年7期)2014-02-06
