
建瓯万木林省级自然保护区苦槠种群种内与种间竞争
2020-03-08 06:06黄加林
武夷学院学报 2020年12期
黄加林
(光泽县寨里林业工作站,福建 光泽 354100)
苦槠(Castanopsis sclerophylla)为壳斗科栲属常绿阔叶乔木树种,栲属植物中分布最北的一个种。苦槠木材较密致、坚韧,富于弹性,是优良的用材树种;树冠浓密,有一定的观赏价值,是良好的园林绿化树种;脱涩后的果实(种仁)制作的苦槠淀粉可加工成苦槠豆腐、苦槠粉丝和苦槠糕等食品,是具有良好发展潜力的栽培树种[1]。由于长期以来人们对天然阔叶林和次生林的过度采伐和破坏,天然林和次生林面积的日益减少,导致天然苦槠资源日渐枯竭[2]。目前有关苦槠的研究报道主要集中在天然林群落结构与植物多样性[2]、苗木培育[3]、栽培技术[4]、果实的药用价值[5]等方面,有关苦槠次生林种间和种内竞争的研究未见报道。
在结构复杂的森林群落中,植物种群的结构和数量的变化受非生物因素(气候、土壤等)和生物因素(种内和种间竞争)的共同制约,其中后者起着至关重要的作用[6]。由于环境资源是有限的,森林群落中的植物种群在生长发育过程中必然因争夺有限的资源而发生种内和种间的竞争,导致群落植物组成、种群结构、植物多样性的动态变化。因此,植物种内和种间竞争是生态学研究的核心问题[7]。竞争指数是反映植物间竞争程度的指标。徐道炜[8]提出的众多的竞争指数模型可分为距离有关的和与距离无关的两大类,其中与距离有关的Hegyi单木竞争指数模型能更好地反映种间和种内竞争程度,而被广泛使用。笔者利用Hegyi单木竞争指数模型对福建万木林自然保护区苦槠群落的种间和种内竞争进行研究,揭示苦槠种群的生态适应机制和动态变化,为苦槠资源的科学保护和可持续利用提供参考资料。
1 试验地概况
试验地位于福建省建瓯市房道镇万木林自然保护区的以苦槠为优势种的次生林群落,北纬27°02.987'~27°03.032',东经117°08.545'~117°08.685',海拔430~460 m左右,属亚热带海洋性季风气候,年均气温18.7℃,年均降水量1 673 mm,年均相对湿度约81%,年均无霜期290 d[2]。
2 研究方法
2.1 外业调查方法
在苦槠次生林群落内连续设立20个10 m×10 m样方,在每块样方中选取乔木层(树高5 m及以上)苦槠为对象木,测定其胸径、树高、冠幅、GPS坐标等指标。根据群落内乔木层大树的冠幅确定竞争木为距离对象木8 m以内的所有乔木树种[8]。以每株对象木为圆心,8 m为半径设置样圆,记录样圆内所有高度5 m及以上的乔灌木树种(竞争木)的树种名称、胸径、树高、冠幅、GPS坐标、与对象木的水平距离等指标。
2.2 竞争指数模型的选择
单木竞争指数模型种类较多,其中Hegyi提出的竞争指数模型使用简便,数据容易获得,在研究中得到较多的应用,本文采用该模型计算竞争指数[7-14]:

式中:CIi为第i株对象木(苦槠)的竞争指数;Di为第i株对象木的胸径;Dj为第j株竞争木(伴生树种或苦槠)的胸径;Lij为对象木i与竞争木j之间的水平距离;n为第i株对象木的竞争木株数;CI为所有对象木(苦槠种群)与竞争木的竞争强度;m为对象木的总株数。CI值越大,表明树种间的竞争越激烈,对象木受到的竞争压力越大。当竞争木全为苦槠时,CIi为第i株对象木的种内竞争指数,CI为苦槠种群的种内竞争强度;当竞争木全为伴生树种时,CIi为第i株对象木的种间竞争指数,CI为苦槠种群的种间竞争强度。
2.3 竞争指数与对象木胸径的关系
树种的竞争能力除了与其生物学、生态学特性和立地条件有关外,还与对象木个体大小有关。将每株对象木的平均竞争指数与其胸径分别用倒数模型和幂函数模型等2个数学模型进行拟合,找出最佳的平均竞争指数与对象木胸径的数学模型:

式中:CI为某对象木与所有竞争木的竞争指数的平均值,D为某对象木胸径,A、B为模型参数。
3 结果与分析
3.1 对象木与竞争木概况
所有样方的对象木(苦槠)有21株,最小胸径16.2 cm,最大胸径83.2 cm,老树较多,没有小径级的小树和幼树,种群处于老化和衰退阶段。对象木30~35 cm和40~45 cm径级的株数最多,均占总株数的19.05%,其他径级的株数占比从大到小依次是:20~25 cm(14.29%)、35~40 cm(9.52%)、45~50 cm(9.52%),15~20、25~30、50~55、55~60、65~70和80~85 cm径级的株数最少,均占总株数的4.76%见表1。

表1 苦槠(对象木)和竞争木的径阶分布Tab.1 Diameter distribution of Castanopsis sclerophylla trees(objective trees)and their competitive trees
竞争木共294株,最小径级平均胸径4.6 cm,最大胸径90.0 cm,由苦槠、罗浮栲、木荷、栲树、薄叶山矾、桂北木姜子、山矾、枫香、闽楠、檵木等38个树种组成。竞争木的径级分布与对象木不同,5~10 cm径级的株数所占的比例最大,高达51.70%,其次是10~15 cm径级,占竞争木总株数的14.97%,该径级以上的竞争木株数总体上随径级的增大而减少。胸径50 cm以上大径级竞争木主要由枫香、苦槠、马尾松、木荷、罗浮栲、栲树等树种组成。
3.2 苦槠种内与种间竞争
苦槠种内竞争强度为8.712,种间竞争强度为27.038,占总竞争强度的75.63%,说明苦槠的竞争主要来自种间,这主要与苦槠径级较大、株数较少、分布较分散有关。
不同径级对象木受到的种内竞争强度均低于种间竞争强度,但种内单株平均竞争指数除了55~60 cm径级外均高于种间竞争指数,这主要是由于苦槠大径级的比例远高于伴生树种。最小径级(15~20 cm)对象木受到的竞争强度最大,种内和种间单株平均竞争指数分别为0.558和0.191,而最大径级(80~85 cm)对象木受到的种内和种间单株平均竞争指数仅分别为0.158和0.020,。苦槠种内和种间竞争强度总体上均有随着径级的增大而减小的趋势,但也有一定的波动,尤其是种间竞争强度有较大幅度的波动(表2)。

表2 苦槠的种内和种间竞争强度Tab.2 Intraspecific and interspecific competition intensity of Castanopsis sclerophylla
虽然苦槠受到种内竞争强度(27.038)低于总的种间竞争强度,但其数值高于其他任何一种伴生树种,见表3。在伴生树种中,苦槠与罗浮栲的竞争指数最大(5.431),其次是木荷(3.487)、栲树(2.267)、薄叶山矾(1.829)、桂北木姜子 (1.744)、山矾 (1.696)、枫香(1.478)、闽楠 (1.109)、檵木 (0.857)、福建山矾(0.813)、椤木石楠 (0.649)、山黄皮 (0.566)、野柿(0.510)、浙江桂(0.453)、米槠(0.409)、树参(0.334)、华杜英(0.334)、栓叶安息香(0.323)、秀丽锥(0.318)、台湾冬青(0.275)、虎皮楠(0.263)、青冈(0.236)、秀丽四照花(0.205)、榕叶冬青(0.178)等。
3.3 苦槠对象木胸径与竞争指数的关系
树种间的竞争强度除了受树种的生态习性、生活幅度和立地条件等多种因素制约外,对象木的个体大小对竞争强度有较大影响。采用倒数模型和幂函数模型拟合苦槠对象木胸径与平均单株竞争指数的关系,结果表明,苦槠种内单株平均竞争强度与对象木胸径有显著的幂函数关系(P=0.039<0.05),与对象木胸径倒数的关系不显著(P=0.152>0.05);苦槠与伴生树种种间单株平均竞争强度与对象木胸径有极显著的幂函数关系(P=0.004<0.01),与对象木胸径的倒数也有极显著的线性相关关系(P=0.008<0.01),但幂函数关系更为密切;苦槠与所有树种的单株平均竞争强度与对象木胸径的倒数有极显著的线性相关关系(P=0.000<0.01),与对象木胸径也有极显著的幂函数关系(P=0.001<0.01),但与对象木胸径的倒数线性相关关系更紧密,见表4、图1。
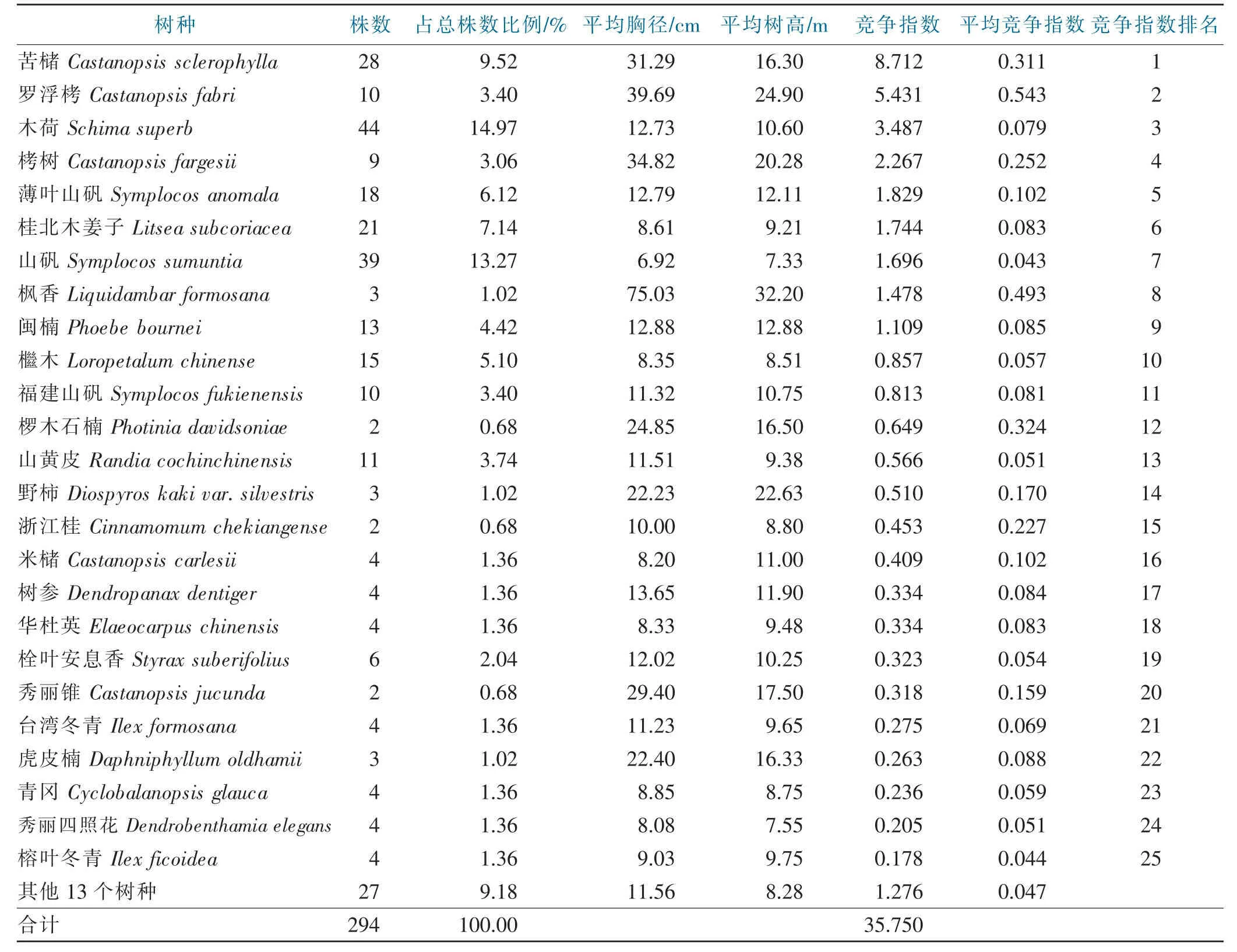
表3 竞争木的种类组成、株数和竞争强度Tab.3 Composition of competitive trees and their competitive indices
从苦槠对象木胸径与竞争指数的关系数学模型可以看出,随着对象木胸径的增加,竞争指数急剧减小,对象木胸径小于40 cm受到的竞争压力较大,胸径40 cm以上的对象木竞争指数较低。
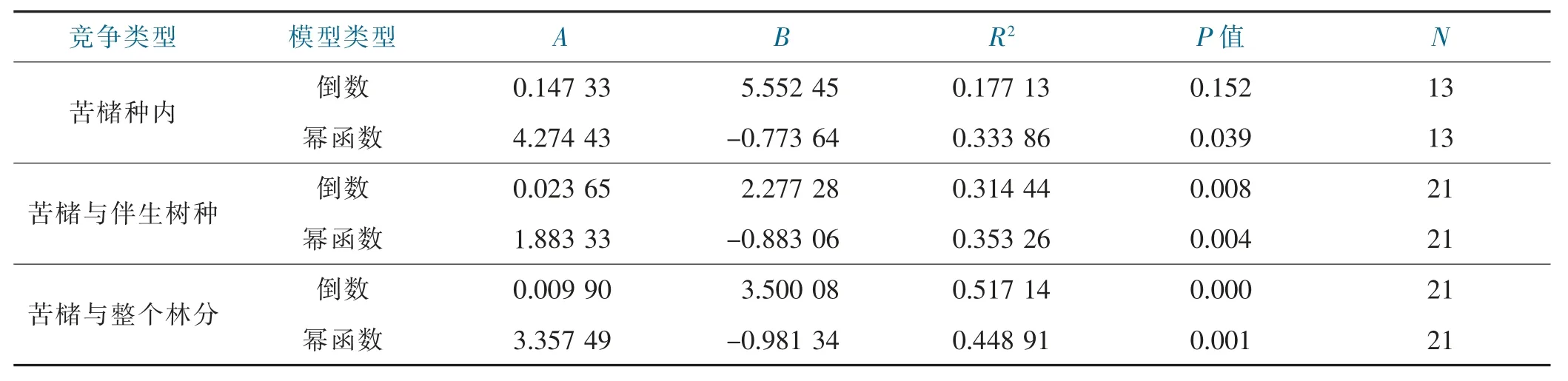
表4 对象木竞争强度与胸径的回归模型参数Tab.4 Parameters of CI-DBH models of object trees

图1 对象木胸径与种内、种间竞争指数与的关系Fig.1 Relationship between DBH of objective trees and intra-and interspecific competitions
4 小结与讨论
众多的研究表明,使用与距离有关的Hegyi单木竞争指数模型对森林群落中林木个体间的竞争进行定量研究,可更准确地度量林木个体间的竞争程度,进而描述和预测种群的动态变化[7-8]。
通过对苦槠群落乔木层21株苦槠对象木和294株竞争木的研究表明:乔木层苦槠种内竞争强度为8.712,高于其受到的任何一种其他伴生树种的种间竞争强度,但低于种间竞争指数总和(27.038),乔木层对象木的竞争压力主要来自种间,这与项小燕等[10]对大别山五针松、曾思齐等[11]对湖南平江青冈栎和李帅锋等[12]对滇西北云南红豆杉的种内和种间关系的研究结果相似,而与黄小波等[13]对云南云龙云南松和王晓霞等[14]对山西太岳山脱皮榆等优势度较大的树种的种内和种间关系的研究结果相反。苦槠在群落中的株数虽少,但分布较为集中,径阶较大,这是苦槠的种内竞争指数高于其受到的任何一种其他伴生树种的种间竞争强度的最大的原因。
在伴生树种中,乔木层罗浮栲的竞争指数最大(5.431),其次是木荷(3.487)。苦槠对象木受到的种内和种间竞争强度的大小顺序为:苦槠>罗浮栲>木荷>栲树>薄叶山矾>桂北木姜子>山矾>枫香>闽楠>檵木>福建山矾>椤木石楠>山黄皮>野柿>浙江桂>米槠>树参>华杜英>栓叶安息香>秀丽锥>台湾冬青>虎皮楠>青冈>秀丽四照花>榕叶冬青等。
树种间的竞争强度与各树种的生物、生态学特性和生态幅度有关,对象木的种间和种内竞争强度还受到自身和竞争木胸径、密度、所处生境等因素的共同影响。福建万木林保护区的次生林的形成已有几百年的历史,在演替初期林下光照充足,喜光的马尾松、枫香等先锋树种竞争力强而成为优势树种,但随着群落的演替,林下光照强度逐渐减弱,先锋树种的幼苗幼树耐阴性比常绿阔叶树差,竞争力低,逐渐被淘汰。苦槠次生林是次生林演替过程中最早出现的常绿阔叶林类型之一[15],苦槠种群缺乏幼苗幼树,主要原因是其幼苗幼树较为喜光,目前的森林环境中林下光照不足,幼苗幼树难以生长发育成小树,再加上且其种子容易被鸟类和啮齿目动物采食,导致能够顺利发育成幼苗的种子数量极少。
苦槠种内和种间单株平均竞争强度随对象木胸径的增大而急剧减小,呈显著的幂函数关系。对象木胸径小于40 cm时受到的竞争压力较大,对象木胸径大于40 cm时受到的竞争强度较低且趋于稳定。一些处于林缘的苦槠由于胸径和冠幅较大,竞争压力小,结果量大,因此,为了解决苦槠天然更新能力差的问题,保护和开发现有的苦槠资源,建议选择生长性状较好、结果量大的苦槠优树进行采果育苗繁殖,大力营造苦槠人工林,加强苦槠天然林资源的保护、开发和合理利用。
猜你喜欢
广西林业科学(2022年4期)2022-11-07
河南师范大学学报(自然科学版)(2022年5期)2022-08-08
中国农业科学(2022年10期)2022-06-28
农业科技与信息(2022年9期)2022-06-18
林业调查规划(2022年3期)2022-06-13
广东蚕业(2021年12期)2022-01-18
园艺与种苗(2021年3期)2021-05-26
临床检验杂志(电子版)(2020年1期)2020-04-03
热带农业科学(2019年8期)2019-09-19
江苏农业科学(2017年6期)2017-05-11
