
桂西南喀斯特山地蚬木叶片性状对微地形变化的响应
2020-02-26 01:04欧芷阳庞世龙申文辉何琴飞
西南农业学报 2020年10期
欧芷阳,庞世龙,何 峰,申文辉,何琴飞,郑 威
(广西壮族自治区林业科学研究院,广西 南宁 530002)
【研究意义】植物功能性状是其在长期适应环境过程中形成的能最大程度利用各种外部资源的形态、结构、生理和物候等属性[1],主要由遗传因素和环境条件共同决定[2],能客观反映植物对环境的适应能力。其中,叶片是植物进行光合作用的主要器官,是植物与外界环境接触面积最大的部位[3],其性状更易于测定,是当前植物功能性状研究的最主要对象[4-6]。从易于测定的植物叶片性状入手,研究植物叶片性状随着环境梯度变化而变化的规律,可预测植物为应对环境变化所采取的适应策略[7-9]。喀斯特地貌结构复杂、地形破碎、生境异质性强,在一定的海拔高差范围内生境类型可产生巨变[10]。蚬木(Excentrodendronhsienmu)是桂西南石灰岩季节性雨林的优势树种,属于国家II级保护植物,因人为干扰及种植经济作物对蚬木分布区域的蚕食,蚬木种群分布面积已极大缩小,保护现状堪忧。因此,分析蚬木叶片性状对微地形变化的响应规律,更有助于理解喀斯特植物的环境适应机制,对桂西南喀斯特山地的植被恢复和生态重建具有重要意义。【前人研究进展】植物的功能性状变异如植物群落的开花格局在大尺度上主要受气候条件(温度和降雨)的影响[11],植物的叶氮浓度随着气温的升高而增加[12],比叶面积随着降水量的增加而增大[13]。Meng等[14]研究我国80个样点温湿梯度下叶片性状的数量变异,发现单位面积叶氮浓度和叶钾浓度随着温度的升高而下降,单位面积叶磷浓度则呈升高趋势。Wright等[15]通过建立叶片能量输入和输出平衡模型,发现白天和夜间叶片与空气的温差是影响叶片大小地理梯度的关键。地形对植物性状的影响较复杂,研究小尺度上地形对植物功能性状的影响,可解释大尺度研究中无法解释的环境过渡效应[16]。其中,不同坡位的叶氮浓度、比叶面积和叶磷浓度等性状存在显著差异[17-19];不同坡向上的叶片功能性状也存在显著差异,一般阳坡优势种较阴坡优势种具有更高的叶δ13C,且在旱季具有较高的钾含量,高水分利用效率和钾素累积是阳坡植物适应其生境的重要手段[20],阴坡的比叶面积较大,而叶干物质含量在阳坡较高[21-22];海拔是影响山地植物生长发育的重要地形因子,植物叶片性状受海拔引起的水热变化影响,在海拔梯度上植物的叶面积、鲜重、干重和比叶面积等性状随着海拔的升高产生显著变化[2-3]。【本研究切入点】近年来已有研究者从种群生态学、群落生态学及抗性生理等方面对蚬木展开了一系列研究[23-25],但对异质性生境下蚬木叶片功能性状的研究鲜见报道。【拟解决的关键问题】测定桂西南喀斯特山地优势树种蚬木的叶片性状,分析微地形变化对其叶片功能性状的影响,为该区域的植被恢复和生态重建提供理论依据。
1 材料与方法
1.1 研究区概况
调查在广西西南部的平果、隆安和武鸣3县(区,东经106°33'~108°37',北纬22°8'~23°51')进行,这些区域属南亚热带季风气候区,年均气温19~22 ℃,年降水量1100~1700 mm,年均相对湿度75 %以上,年均日照时数1597~1660 h,降雨多集中于5-9月,雨、热同期。调查区域喀斯特地貌典型,基岩大面积裸露、土层浅薄,土壤以黑色、棕色和红色石灰土为主。石灰岩季节性雨林是本区域的代表性植被[26],广泛分布于桂西南喀斯特山地,群落建群种为蚬木,圆叶乌桕(Sapiumrotundifolium)和苹婆(Sterculianobilis)为乔木层常见伴生种。群落下层常分布有广西密花树(Rapaneakwangsiensis)、岩柿(Diospyrosdumetorum)、石山棕(Guihaiaargyrata)、米扬噎(Streblustonkinensis)和葡萄叶艾麻(Laporteaviolacea)等灌木物种,草本层稀疏,以岭南铁角蕨(Aspleniumsampsoni)、石生铁角蕨(A.saxicola)和假鞭叶铁线蕨(Adiantummalesianum)等较常见。
1.2 样品采集及环境因子调查
基于前期的野外调查结果,于2017年11-12月在桂西南平果县马头镇、隆安县布泉镇和南宁市武鸣区陇均—三十六弄自然保护区的典型喀斯特山地进行蚬木叶片采集。其中,在平果和隆安县各采集9个样点,在武鸣区采集13个样点,共31个样点。叶片采集时,在每个20 m×20 m样方内各选择5株生长良好、病虫害少的蚬木,在每株的冠层上方4个方向各采集生长良好且无遮光的成熟叶片12片,用水湿润后迅速装入自封袋,置于样品箱中带回室内进行相关指标测定。同时用手持式罗盘仪记录采集点的经纬度、海拔、坡度、坡位和坡向等因子。样品采集点数量和样本数量详见表1。
1.3 测定指标及方法
测定指标包括叶片鲜重、干重、叶干物质含量(LDMC)、叶片厚度、叶面积、比叶面积(SLA)、相对叶绿素含量、叶氮浓度(LNC)和叶磷浓度(LPC)等9个指标。
1.3.1 叶干物质含量测定 将叶片从自封袋内取出,用吸水纸将叶片表面多余水分吸干,用电子天平(精度0.0001 g)称取每片叶片的鲜重。将称重后的叶片置于80 ℃烘箱烘干至恒重,用电子天平(精度0.0001 g)称取每片叶片的干重。LDMC(g/cm2)=叶干重/叶鲜重。
1.3.2 叶片厚度测定 沿叶片主脉一侧均匀选3个点,用游标卡尺(精度0.02 mm)测定各点的厚度,取3个点厚度值的平均值作为叶片厚度(mm)。
1.3.3 比叶面积测定 采用CI-203手持式叶面积仪进行叶面积扫描,并计算比叶面积。SLA(mm2/mg)=叶面积/叶干重。
1.3.4 相对叶绿素含量测定 采用CCM-200便携式植物叶绿素仪测定叶片的相对叶绿素含量,测量时沿主脉一侧均匀选3个点,以这3个点测量值的平均值作为叶片相对叶绿素含量(%)。
1.3.5 叶氮浓度和叶磷浓度测定 参考鲍士旦[27]的方法采用凯氏定氮法测定叶片全氮含量,采用钼锑抗比色法测定叶片全磷含量,计算叶氮浓度和叶磷浓度。LNC(mg/g)=叶片全氮含量/叶片干重,LPC(mg/g)=叶片全磷含量/叶片干重。
1.4 统计分析
海拔和坡度采用实测数据进行分析;对坡位和坡向进行赋值,以1代表下坡位,2代表中坡位,3代表上坡位;坡向数据以顺时针方向旋转的角度表示,以每90°为一个区间划分等级:1表示北坡(315°~45°),2表示东坡(45°~135°),3表示南坡(135°~225°),4表示西坡(225°~315°);采用K-S法检验样品数据是否符合正态分布,不符合正态分布的数据经倒数转换后再进行数据分析。采用STATISTICA 8.0对叶功能性状和地形因子进行相关分析和Kruskal-Wallis检验,以SPSS 19.0进行Duncan's新复极差法多重比较,以DPS对采集点地形信息和蚬木叶性状信息进行灰色关联度分析。
2 结果与分析
2.1 叶片性状间的相互关系
由表1可知,蚬木叶片性状的变异系数排序为比叶面积>叶干重>叶磷浓度>叶鲜重>叶面积>叶干物质含量>叶片厚度=叶氮浓度>相对叶绿素含量,表明比叶面积变化较大,而相对叶绿素含量变化较稳定。Pearson相关分析结果(表2)显示,叶面积与鲜重呈极显著正相关(P<0.01,下同),与干重呈显著正相关(P<0.05,下同),与叶氮浓度呈显著负相关,即面积较大的叶片其鲜重和干重相对较重;鲜重和干重与叶氮浓度呈极显著负相关;比叶面积与叶干重和叶干物质含量呈极显著负相关,与叶氮浓度呈极显著正相关;叶干物质含量与叶片厚度呈显著负相关,与叶氮浓度呈极显著负相关,而叶片厚度与叶鲜重呈显著正相关;相对叶绿素含量和叶磷浓度与其他叶性状间相关性不显著(P>0.05,下同)。可见,蚬木的叶鲜重随着叶厚度增加而增加,但叶干物质含量下降。
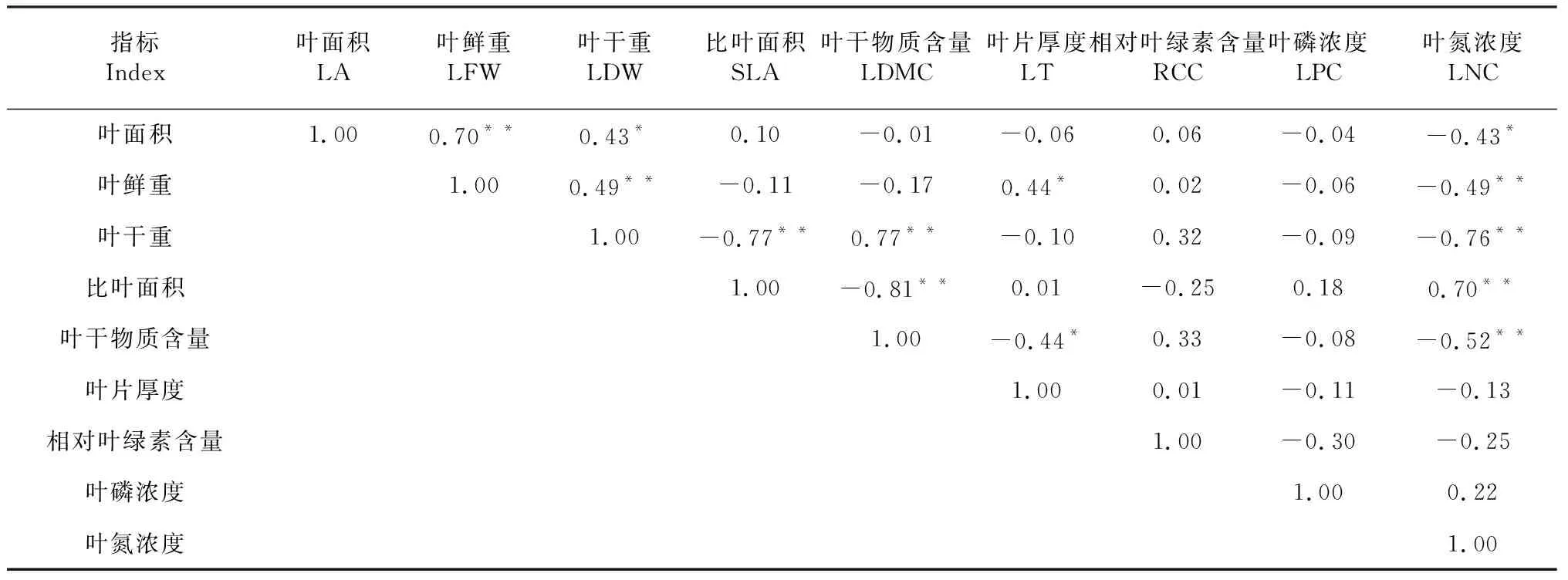
表2 蚬木叶片性状间的相关系数矩阵
2.2 蚬木叶片性状与地形因子的相关性
2.2.1 与海拔和坡度的关系 由表3可知,蚬木叶干物质含量与海拔呈显著负相关,叶片厚度与海拔呈极显著正相关;叶鲜重、比叶面积和叶面积与海拔呈正相关,但相关性不显著;叶干重、相对叶绿素含量、叶磷浓度和叶氮浓度与海拔呈负相关,但相关性均未达显著水平。叶面积、叶片鲜重和干重、比叶面积及叶干物质含量与坡度呈负相关,叶片厚度和相对叶绿素含量与坡度呈正相关,但相关性均未达显著水平;叶氮浓度与坡度间未表现出明显的相关性,叶磷浓度与坡度呈显著负相关。可见,海拔显著影响蚬木叶干物质含量及叶片厚度,坡度显著影响蚬木叶磷浓度的积累。
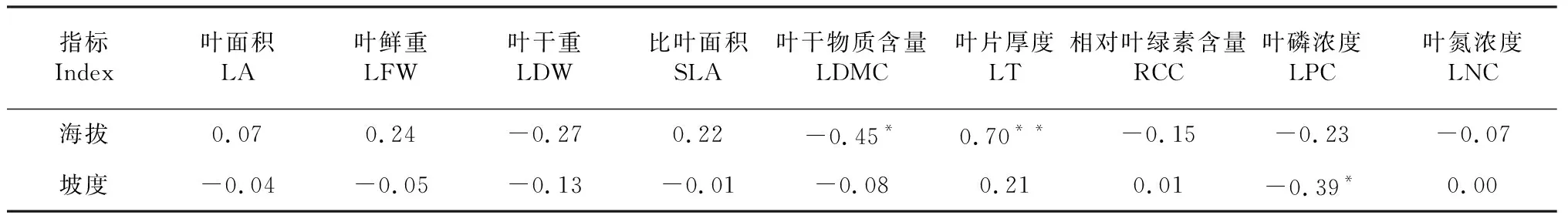
表3 蚬木叶片性状与坡度和海拔的相关性分析结果
2.2.2 与坡位的关系 Kruskal-Wallis检验结果显示,不同坡位上的蚬木叶片鲜重间(H=6.329,P=0.042<0.05)和厚度间(H=6.153,P=0.046<0.05)差异显著(表4)。多重比较结果显示,上坡位的叶鲜重和叶片厚度显著高于下坡位,与中坡位间差异不显著,其随着坡位的变化趋势与随着海拔的升高而增加的趋势一致;上坡位的比叶面积和叶磷浓度显著高于中坡位,中坡位的叶干物质含量显著高于上坡位;不同坡位蚬木叶片的叶面积间、干重间、相对叶绿素含量间和叶氮浓度间差异不显著。表明坡位显著影响蚬木叶鲜重和叶片厚度,在海拔相对较高的上坡位,叶片厚度的增加有利于增强蚬木对水分等资源的保留能力。

表4 不同坡位间的蚬木叶片性状比较
2.2.3 与坡向的关系 Kruskal-Wallis检验结果(表5)显示,不同坡向蚬木叶干重间(H=8.242,P=0.041<0.05)、比叶面积间(H=9.563,P=0.023<0.05)、相对叶绿素含量间(H=8.242,P=0.041<0.05)和叶氮浓度间(H=7.834,P=0.049<0.05)存在显著差异,叶干物质含量间(H=12.339,P=0.006<0.01)存在极显著差异。多重比较结果显示,西坡的叶干重显著高于北坡和东坡,高于南坡但差异不显著;东坡和南坡的比叶面积显著高于西坡,高于北坡但差异不显著;西坡的叶干物质含量显著高于其他3个坡向;西坡的相对叶绿素含量显著高于东坡和南坡,高于北坡但差异不显著;东坡的叶氮浓度显著高于西坡,高于南坡和北坡但差异不显著;不同坡向间蚬木叶片的叶面积、叶鲜重和叶磷浓度无显著差异。表明不同坡向间蚬木叶干重、比叶面积、相对叶绿素含量等性状差异显著,体现了植物对微生境下资源变化的响应。
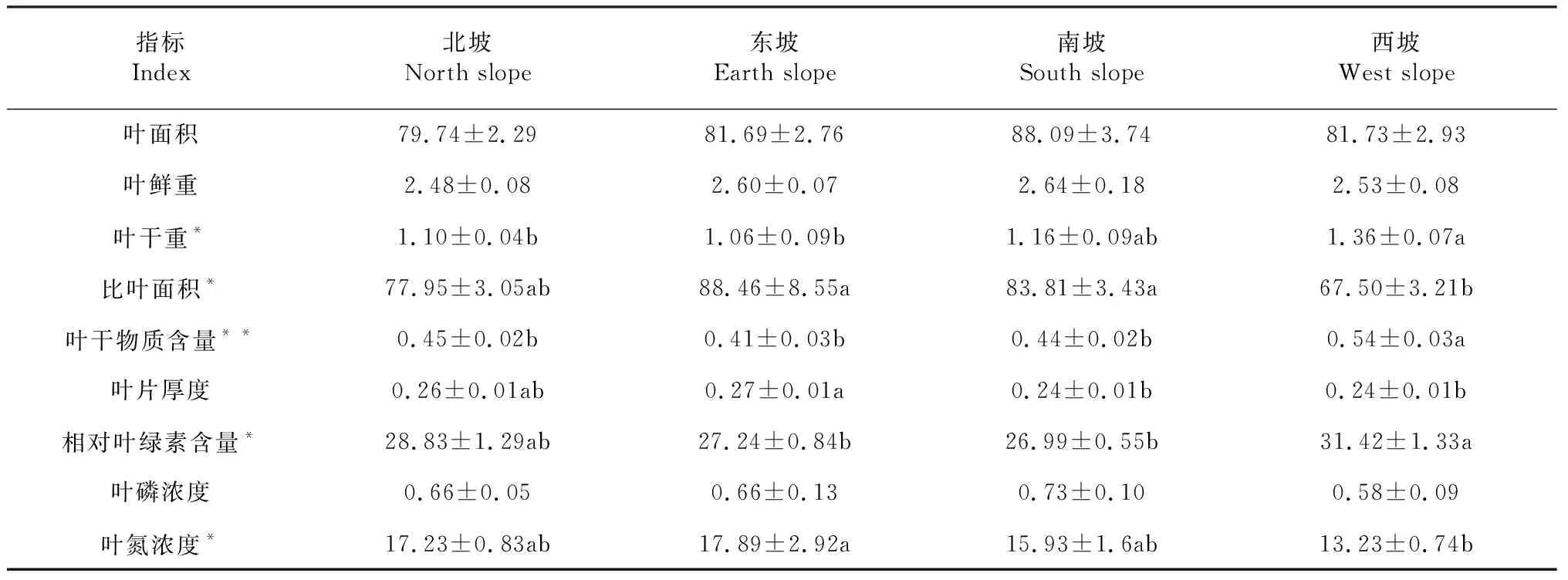
表5 不同坡向间的蚬木叶片性状比较
2.3 蚬木叶片性状与地形因子间的灰色关联度分析
以蚬木叶片的叶面积等性状为因变量,以海拔、坡度、坡位和坡向4个地形因子为自变量进行灰色关联度分析,结果(表6)表明,影响叶面积、鲜重、干重、比叶面积、叶干物质含量、叶片厚度和相对叶绿素含量的地形因子排序为海拔>坡度>坡位>坡向;影响叶磷浓度的地形因子排序为海拔>坡位>坡度>坡向,影响叶氮浓度的地形因子排序为坡度>海拔>坡位>坡向。说明影响叶氮浓度的首要地形因子为坡度,影响其余叶性状的首要地形因子为海拔,而坡向对蚬木叶片各性状的影响均最小。
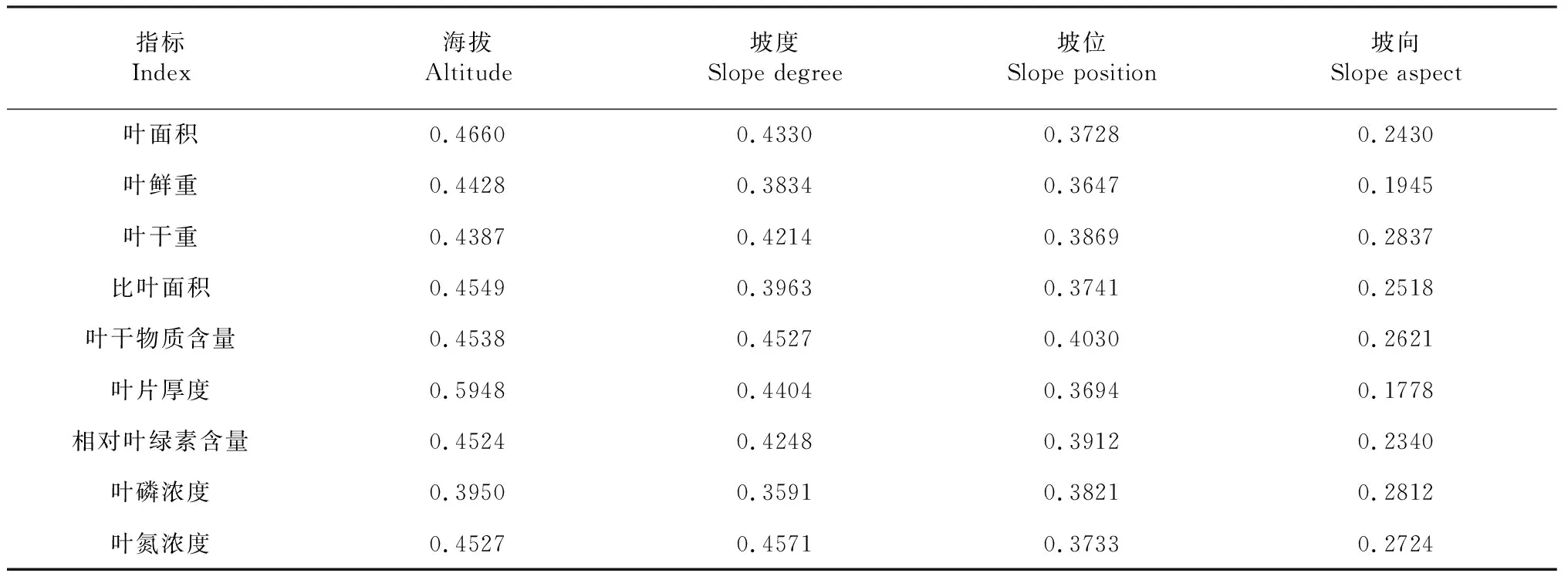
表6 蚬木叶片性状与地形因子的灰色关联度分析结果
3 讨 论
随着海拔的升高,气温下降、年降水量增加、光合辐射增强[28-30],植物的分布及其结构和功能性状也产生显著变化[31-32]。本研究中,影响蚬木叶片大部分性状的首要地形因子为海拔,其中叶干物质含量与比叶面积呈极显著负相关,叶干物质含量随着海拔的升高显著下降,叶片厚度随着海拔的升高极显著增加,与张慧文等[33]研究得到高比叶重(低比叶面积)植物对所获取资源具有较好的保留能力的结果一致。本研究还发现,在海拔相对较高地段蚬木的叶片厚度较厚、叶干物质含量较低,与盘远方等[22]、孙雪娇等[30]、罗璐等[34]的研究结果一致,说明在海拔相对较高且坡度较大的地段,光照条件虽好但土壤水分流失较快,叶片厚度的极显著增加可增强植物的资源保留能力,减少体内水分和能量消耗。可见,蚬木的叶干物质含量和叶片厚度随着海拔梯度的变化主要体现其对光照资源和土壤水分变化的适应。
坡度从垂直结构上影响土壤水肥的空间分布,坡度越大的地段,土壤的水肥越容易流失。蚬木的叶片厚度和相对叶绿素含量随着坡度的增加而增加,叶面积、鲜重和干重、比叶面积及叶干物质含量等呈下降趋势,叶磷浓度与坡度间呈显著负相关,与刘贵峰等[5]、Han等[35]的研究结果相似,与陈晨等[3]、党晶晶等[36]研究认为坡度大的地段土壤水肥匮乏、植物需要将更多的资源投入到地下部分生长、对叶片建成的投资减少,导致叶面积和叶重量等性状随着坡度的增加而减小的观点一致。本研究还发现,坡位对蚬木叶片性状的影响规律与对海拔的影响规律一致,其中上坡位蚬木叶片的比叶面积最大,叶片厚度由下坡位至上坡位逐渐增加,上坡位的叶片厚度显著高于下坡位。
坡向影响光、温、水、风等生态因子的组合,间接影响植物的生长发育。已有研究表明,不同坡向环境因子存在显著差异,植物的性状也存在显著差异[22,37]。本研究结果与上述研究结果相似,不同坡向间蚬木的叶干重、比叶面积和相对叶绿素含量差异显著,体现了蚬木对环境资源变化的响应。其中,阴坡面(北坡)蚬木的叶面积最小,光照最充足的南坡叶面积和叶鲜重最大,与高辉[38]研究认为蚬木作为阳生树种,幼龄前期需荫蔽、之后逐渐喜光,过于荫蔽的环境影响蚬木生长发育的观点一致;生长在环境条件最适宜东坡蚬木叶片的厚度最厚、比叶面积和叶氮浓度最大、叶干物质含量最低,但生长于西坡蚬木的叶干重和叶干物质含量最大或最高,比叶面积最小;与资源相对丰富的东坡面相比,西坡微生境更为高温干旱,叶片的高叶干物质含量、低比叶面积性状有利于蚬木对所获取资源的利用。
4 结 论
不同地形(海拔、坡度、坡位和坡向)条件下的蚬木叶功能性状存在差异,是蚬木适应喀斯特高度异质性微生境的策略。
猜你喜欢
农学学报(2022年8期)2022-11-16
军事文摘(2022年14期)2022-08-26
散文诗(2021年22期)2022-01-12
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02
新疆农业科学(2020年1期)2020-02-14
环球时报(2019-05-23)2019-05-23
安徽农学通报(2017年11期)2017-06-23
绿色科技(2017年1期)2017-03-01
湖北农业科学(2014年16期)2014-10-20
现代农业科技(2009年1期)2009-04-09
