
狭叶柴胡β-amyrin合成酶基因的克隆及其蛋白的生物信息学分析
2020-02-25 08:30:36刘艳玲向凤宁
西南农业学报 2020年11期
刘艳玲,向凤宁
(1.德州学院医药与护理学院,山东 德州 253023;2.山东大学生命科学院植物细胞工程与种质创新教育部重点实验室,山东 青岛 266237)
【研究意义】通过克隆植物代谢通路上某一关键酶基因并对其蛋白进行生物信息学分析可为该代谢途径的调控机理提供一定的理论依据。【前人研究进展】狭叶柴胡(Bupleurumscorzonerifolium)系伞形科(Umbelliferae)柴胡属(Bupleurum)多年生草本植物,主要分布于我国东北、华北、西北、江苏、四川等地[1-2]。《中国药典》2015年版中规定狭叶柴胡的干燥根为药材柴胡[3],主要成分为柴胡皂甙、甾醇、挥发油和多糖,其中柴胡皂甙类化合物含量较高,柴胡皂苷d (SSd) 的药理活性最强[4]。现代药理学研究表明狭叶柴胡具有镇痛、镇静、抗病毒、调节免疫、降血脂等功效[5-6],已广泛应用于肾病、肝纤维化、肿瘤等疾病的治疗,并取得了显著疗效,其中尤以保肝作用效果最佳,这也与传统中医记载柴胡疏肝解郁功效一致[7]。柴胡皂甙d结构为五环三萜齐墩果烷型,β-amyrin 合成酶是齐墩果酸合成的关键酶之一,属于氧化角鲨烯环化酶(OSCs)家族[8-10]。目前已从人参[11]、豌豆[12]、甘草[13]、白桦[14]、拟南芥[15]等多种植物中成功克隆得到编码β-amyrin合成酶的cDNA序列,但尚未见狭叶柴胡 β-amyrin合成酶基因克隆及其生物信息学分析的报道。【本研究切入点】本研究中,首先克隆获得狭叶柴胡 β-amyrin 合成酶基因的全长序列,然后采用生物信息学方法,对该基因结构、蛋白质理化特性、空间结构、磷酸化位点、亚细胞定位等进行预测和分析。【拟解决的关键问题】有望为齐墩果烷型代谢产物及代谢途径的深入研究奠定基础。
1 材料与方法
1.1 供试材料
实验材料狭叶柴胡采自内蒙古。
1.2 试剂
DNA聚合酶、RNA酶抑制剂、Olig(dT)12-18、5×M-MLV reverse transcriptase Buffer、10 mM each dNTPs、琼脂糖凝胶DNA回收试剂盒、DNaseI(4 U/μl)、限制性内切酶、T4DNA 连接酶、Solution I、pMD18-T载体等购自TaKaRa公司;TRIZOL、DEPC、琼脂糖、LB培养基等购自上海生物工程技术服务有限公司。
1.3 试验方法
1.3.1 BsAS全长序列扩增 (1) BsAS基因中间片段的扩增。正向引物 P1: 5’-TGGCTTTCGATA(T)CTTGGA-3’;反向引物 P2:5′-CCACCG(A)TTTTTG(A)CTCTGTA-3’。
(2)BsAS基因5’端片段的扩增。采用20 μl PCR体系,首先P3与P4进行扩增,将PCR产物取0.2 μl为模板进行P4与P6的扩增,再将二次PCR产物取0.2 μl为模板进行P5与P6的扩增。
5′半套式PCR反向引物。P3: 5′-TGACGGCGTCATACATTTTC-3′;P4: 5′-CTCAACCCAACAAGCAA GC-3′;P5: 5′-TTTCATCTTCATAATGTATATG-3′。正向引物。P6: 5′-ATGTGGARGCTDAAGATHG-3′。
(3)BsAS基因3’端片段的扩增。采用3’RACE(Rapid Amplification of cDNA Ends)方法进行扩增,参照Clonetech, SMART RACE cDNA Amplification Kit操作指南进行。正向引物为P7:TCTTACTATCATTACAGAGCA;反向引物为3’RACE试剂盒中自带。
(4)BsAS全长基因的扩增。正向引物P8:5′-ATCCGAAAGGAGACTATTTCA AG-3′;正向引物P9:5′- TTAAAGAGTAGTGGAAGGCA A-3′;扩增程序:94 ℃ 5 min;94 ℃ 1 min, 60 ℃ 2.5 min,72 ℃ 1 min,35个循环;72 ℃ 10 min;10 ℃保存。
以上引物均由上海生物工程技术服务有限公司合成,测序由博尚生物公司完成,核苷酸及蛋白质序列比对分析利用DNAMAN软件中的比对程序进行。
1.3.2 BsAS序列生物信息学分析 使用ExPASy的ProParam在线数据库对狭叶柴胡BsAS序列进行理化性质的预测分析;应用TMHMM (http://www.cbs.dtu.dk/services/TMHMM/)在线工具对狭叶柴胡BsAS序列进行跨膜结构的预测分析;利用PSORT Ⅱ软件对狭叶柴胡BsAS蛋白进行亚细胞定位;利用Conserved Domains Search 网站(https://www.ncbi.nlm.nih.gov/Structure/ cdd/wrpsb.cgi)对狭叶柴胡BsAS氨基酸进行保守域分析;采用 SOPMA 在线工具预测狭叶柴胡BsAS蛋白质二级结构;使用Swi-ssModel 网站(https://www.swissmodel.expasy.org/)中的在线数据库构建狭叶柴胡BsAS 蛋白质三级结构;应用NetPhos3.1 server在线软件对狭叶柴胡BsAS序列的磷酸化位点进行预测应用MEGA6.0对狭叶柴胡进行系统进化树的分析。
2 结果与分析
2.1 狭叶柴胡中β-amyrin合成酶基因BsAS的克隆
首先从Genebank中下载甘草、豌豆、人参、白桦的β-amyrin合成酶基因序列,设计简并引物P1、P2,在狭叶柴胡cDNA中进行扩增,获得约900 bp的一条片段,经测序、序列比对确定为β-amyrin合成酶基因片段。然后5’片段使用半套式PCR方法,3’片段使用RACE方法进行扩增,最后通过引物P8和P9进行基因全长的克隆(图1),经BLAST分析,结果表明,该基因编码β-amyrin合成酶,全长2289 bp,共编码762 aa,命名为BsAS(β-amyrin synthase gene)。BsAS的核苷酸及氨基酸序列见图2。

1:中间片段扩增,2:5’片段扩增,3: 3’片段扩增,BsAS:全长cDNA扩增1: Intermediate fragment amplification,2: 5’fragment amplification,3: 3’fragment amplification,BsAS:Full-length cDNA amplification

图2 BsAs核苷酸及氨基酸序列Fig.2 Sequence of BsAs nucletide and aiminoacids
2.2 狭叶柴胡BsAS基因生物信息学分析
2.2.1 BsAS保守序列分析 研究表明,氧化鲨烯环化酶家族蛋白具有高度保守的DCTAE、QW和MWCYCR 3个序列。通过序列分析,发现BsAS中含有4个拷贝的QW序列、1个DCTAE序列和1个MWCYCR序列(图3)。

a.QW阅读框架;b.PCTAE阅读框架;c.MWCYCR阅读框架
利用Conserved Domains Search 网站(https://www.ncbi.nlm.nih.gov/Structure/ cdd/wrpsb.cgi)对狭叶柴胡BsAS氨基酸进行保守域分析,发现狭叶柴胡BsAS序列的1~758 位氨基酸是它的功能区域,注释为Camelliol C合酶,该酶属于氧化鲨烯环化酶家族 (图4)。

图4 狭叶柴胡BsAS氨基酸保守区分析Fig.4 Conserved domain prediction of B. scoronerifolium BsAS amino acid
2.2.2 狭叶柴胡BsAS理化性质分析 使用ExPASy的ProParam在线数据库对狭叶柴胡BsAS序列进行理化性质的预测分析。推测BsAS序列的分子式为C3955H5944N1046O1112S48,分子量为87.47592 kDa, pI为6.04,带负电残基(Asp + Glu)为90,带正电残基(Arg + Lys) 为75,蛋白的不稳定系数为50.25,属于不稳定蛋白;脂肪系数为74.79,BsAS蛋白的亲水性平均系数为-0.357,为亲水性蛋白。
使用 ExPASy 的在线分析工具 Protscale 对狭叶柴胡BsAS 蛋白的亲疏水性做进一步预测(图5),结果表明:狭叶柴胡BsAS氨基酸多肽链的亲水性最小值为-2.811,最大值为2.644;BsAS中亲水性氨基酸所占比例较大,疏水性氨基酸所占比例较小,其结果与ProParam软件预测相一致。

图5 狭叶柴胡BsAS的疏水性分析Fig.5 Hydrophobicity analysis of B. scorzonerifolium BsAS
2.2.3 狭叶柴胡BsAS跨膜区、亚细胞定位分析 应用TMHMM (http://www.cbs.dtu.dk/services/TMHMM/)在线工具对狭叶柴胡BsAS序列进行跨膜结构的预测分析,结果显示该蛋白没有跨膜螺旋区(图6)。

图6 狭叶柴胡BsAS序列跨膜区域预测Fig.6 Transmembrane domain prediction of B. scorzonerifolium BsAS
利用PSORT Ⅱ软件对狭叶柴胡BsAS蛋白进行亚细胞定位,发现狭叶柴胡BsAS蛋白有39.1 %位于细胞质中,26.1 %位于线粒体上,17.4 %位于细胞核内,4.3 %位于内质网中,4.3 %位于分泌系统的囊泡中,4.3 %位于过氧化体,其余4.3 %位于高尔基体,表明狭叶柴胡BsAS蛋白主要存在于细胞质中。
2.2.4 狭叶柴胡BsAS二级结构预测 采用SOPMA在线工具预测狭叶柴胡BsAS蛋白质二级结构(图7),狭叶柴胡BsAS蛋白二级结构的氨基酸序列中α-螺旋(Hh)为45.14 %,β-转角(Tt)为7.35 %,无规卷曲(Cc)为 34.38 %,延伸链(Ee)为13.12 %。从组成数据可以看出,该蛋白二级结构的主要元件为α-螺旋和无规则卷曲,β-转角只在局部出现。

图7 狭叶柴胡BsAS二级结构预测Fig.7 Prediction of secondary structure of B. scoronerifolium BsAS
2.2.5 狭叶柴胡BsAS三级结构预测 使用Swi-ssModel 网站(https://www.swissmodel.expasy.org/)中的在线数据库构建狭叶柴胡BsAS 蛋白质三级结构(图8),该模型的全球性模型质量估测(global model quality estimation, GMQE)得分为0.71,在0~1之间,较接近1,由此确定狭叶柴胡BsAS三级结构建模结果可靠。
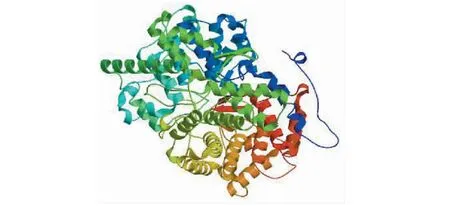
图8 狭叶柴胡BsAS三级结构预测Fig.8 Prediction of tertiary structure of B. scoronerifolium BsAS
2.2.6 狭叶柴胡BsAS磷酸化位点预测 应用NetPhos3.1 server在线软件对狭叶柴胡BsAS序列的磷酸化位点进行预测(图9,表1),结果发现该序列有多个磷酸化位点,但仅有23个丝氨酸、14个苏氨酸、16 个酪氨酸的磷酸化能力超过了阈值,表明狭叶柴胡BsAS只有53个磷酸化位点。其主要的磷酸化位点是蛋白激酶C(PKC)位点、酪蛋白激酶П(CKП)位点和细胞周期蛋白质依赖性激酶(cdc2)位点。

表1 狭叶柴胡BsAS 磷酸化位点列表
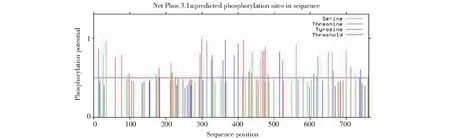
图9 狭叶柴胡BsAS磷酸化位点预测Fig.9 Phosphorylation sites prediction of B. scoronerifolium BsAS
3 讨 论
通过对BsAS基因序列分析发现,该基因含有氧化鲨烯家族特有的3个保守基序DCTAE、QW和MWCYCR。序列DCTAE与底物结合有关;QW序列带负电荷,在 2,3-氧化鲨烯的环化过程中作用于正电荷中间体,达到稳固蛋白质结构的作用;MWCRCY 序列中的色氨酸(W)参与调控三萜化合物碳骨架的生物合成。
通过生物信息在线工具预测狭叶柴胡BsAS 蛋白是亲水蛋白,无跨膜螺旋区,主要在细胞质内发挥生物学作用。蛋白质的磷酸化修饰与多种生物学过程密切相关,20世纪50年代以来一直被生物学家看作是一种动态的生物调节过程。狭叶柴胡BsAS蛋白质含有 53个磷酸化位点,其中最多的磷酸位点是丝氨酸/苏氨酸蛋白激酶C(PKC)。在过去30多年间,PKC家族已被广泛研究,一致认为PKC与多种细胞过程有关,包括肿瘤的形成。研究表明,PKC与肿瘤产生的佛波醇酯相关,其受体会结合并激活PKC,从而导致肿瘤细胞生长加速。因此,医学上一直将PKC作为抑制肿瘤的靶目标[16]。直到2015年,Antal推翻了该理论,认为PKC不但不会产生肿瘤,反而会抑制肿瘤的生长。他首先在结肠癌细胞的基因组中校正了一个功能缺失的PKC突变,然后发现该PKC能抑制小鼠肿瘤的生长,而当PKC失效时,致癌信号增加,肿瘤生长加速,但长期反复处理细胞,PKC长期高表达,则可能导致细胞内部的反馈抑制效应,导致PKC的突变或降解,从而促进了肿瘤的发生[17]。本研究中,狭叶柴胡BsAS蛋白的PKC磷酸化位点较多,可通过磷酸化激活PKC的活性,从而抑制肿瘤产生与发展,这也与柴胡具有抗肿瘤的性质相一致。
4 结 论
本研究通过基因克隆获得了长度2289 bp的β-amyrin合成酶基因,其蛋白命名为BsAS,含有氧化鲨烯环化酶家族所特有的高度保守的DCTAE、QW和MWCYCR开放阅读框架;BsAS无跨膜区,为亲水性蛋白,定位于细胞质中;其二级结构主要是α-螺旋和无规卷曲,三级结构预测结果可信;共有 53个磷酸化位点,以丝氨酸磷酸化位点最多。BsAS全长基因的克隆和序列分析为狭叶柴胡BsAS基因结构与功能研究提供了基础,并可为研究狭叶柴胡三萜合成通路的调节方式提供新的认识。
猜你喜欢
环球时报(2022-09-20)2022-09-20 15:18:57
今日农业(2020年24期)2020-12-15 16:16:00
河北果树(2020年4期)2020-11-26 06:05:18
天津医科大学学报(2019年6期)2019-08-13 07:04:42
中成药(2018年10期)2018-10-26 03:41:02
中成药(2017年6期)2017-06-13 07:30:35
兽医导刊(2016年12期)2016-05-17 03:51:50
兽医导刊(2016年12期)2016-05-17 03:51:46
安徽医科大学学报(2015年9期)2015-12-16 11:09:42
现代检验医学杂志(2015年4期)2015-02-06 02:02:06
