
巴氏杀菌婴儿配方乳对大鼠T细胞亚群及其 血液免疫相关因子的影响
2020-02-08 14:49崔东影席恩泽徐境含李梦寒吴禹昊许晓曦
食品科学 2020年1期
崔东影,席恩泽,徐境含,盛 雪,李梦寒,刘 洋,吴禹昊,许晓曦
(东北农业大学食品学院,黑龙江 哈尔滨 150030)
目前,由于我国母乳数据库还不完善,婴儿配方乳粉虽在营养成分上接近母乳,但免疫活性方面无法达到母乳水平[1],因而最大程度地保持免疫蛋白在婴儿配方食品中的生物活性功效成为乳品生产行业的关键技术[2]。强度高于巴氏杀菌的热处理工艺技术会引起牛乳感官品质下降和营养成分的损失[3-4],不仅降低了乳品质量,也可能会由于一些有害物质的产生导致产品不安全[5-6]。已有的婴儿配方乳粉为安全性考虑均采用高温杀菌,造成了蛋白质结构的改变[7],一定程度上诱导了美拉德反应中间产物的形成,降低了婴幼儿配方食品的消化吸收率[8],由于蛋白质参与矿物质吸收,热损伤的蛋白质可能会影响金属离子的生物利用度[9]。有资料显示,婴幼儿乳粉在冲调过程中容易产生微生物污染的风险,尤其是阪崎肠杆菌对早产儿和低体质量儿会造成极大的危害[10]。婴儿配方乳作为一种乳基液态配方食品,和其他婴儿配方食品相比,具有安全、营养、哺喂方便等特点[11]。与市面现有的经超高温灭菌的婴儿配方产品相比,巴氏杀菌最大程度地保留了鲜牛乳及添加物质的免疫活性和天然营养。目前婴儿配方乳粉中添加的乳铁蛋白具有增强机体免疫、抗菌等生物活性[12-13],但以往研究只停留在不同热处理条件对其结构影响变化阶段,有学者曾将不同热处理条件下的乳铁蛋白及乳铁蛋白素对脾细胞的免疫促进作用[14]以及对癌细胞的促凋亡等功能性[15-16]进行了探讨。但至今针对乳铁蛋白在动物及婴儿体内的生物活性,尤其是对免疫活性的探究仍为空白。本研究利用动物喂养实验将实验室自制巴氏杀菌婴儿配方乳与市售常温婴儿配方乳产品进行对比,用以探究两类不同热处理产品在大鼠生长及血液免疫指标方面的差异性,以期为研制出适合中国婴儿生长发育需要的婴儿配方乳提供科学依据。
1 材料与方法
1.1 动物、材料与试剂
S D 大鼠(雄性、4 周龄、S P F 级、体质量90~110 g),购于北京维通利华实验动物技术有限公司,生产许可证号为SCXK(京)2016-0006。
牛乳、脱盐乳清粉、浓缩乳清蛋白粉 北京银河乳业有限公司;乳糖、二十二碳六烯酸(docosahexeniic acid,DHA)、花生四烯酸(arachidonic acid,ARA) 罗盖特生物营养品有限公司;低聚果糖、低聚半乳糖 保龄宝宝生物有限公司;牛磺酸、核苷酸、左旋肉碱、肌醇、叶黄素、复配维生素、复配矿物质 北京金康普食品科技有限公司;乳铁蛋白、乳白蛋白 天津银河伟业进出口有限公司;大豆油、玉米油、葵花籽油 哈尔滨九三油脂有限公司;结构脂肪 益海嘉里有限公司; 卡拉胶、单甘酯 连云港友进食品添加剂技术开发有限 公司;以上材料均用于实验室自制巴氏杀菌婴儿配方乳。
白细胞介素-2(interleukin 2,IL-2)、IL-4、干扰素-γ(interferon-γ,IFN-γ)、肿瘤坏死因子-α(tumor necrosis factor-α,TNF-α)酶联免疫吸附测定(enzyme-linked immunosorbent assay,ELISA)试剂盒、溶菌酶试剂盒 南京建成生物工程研究所;CD3-FITC、CD4-PE、 CD8a-PerCP单克隆抗体 杭州联科生物技术股份有限公司;肝素钠 上海惠世生化试剂有限公司;磷酸盐缓冲液(phosphate buffered saline,PBS) 北京Biotopped 公司。
1.2 仪器与设备
HH-4数显恒温水浴锅 常州丹瑞实验仪器设备有限公司;JJ高压均质机 廊坊市盛通机械有限公司; HZ实验型超高温杀菌机 上海辉展实验设备有限公司;ACS30型电子秤 浙江风华科技有限公司; HBS-1096A酶标分析仪 南京德铁实验设备有限公司;755B紫外-可见分光光度计 上海精密仪器公司;3K15高速冷冻离心机 德国Sigma公司;DHP-9162电热恒温培养箱 上海一恒科技有限公司。
1.3 方法
1.3.1 实验动物的处理和分组
选择雄性S D 大鼠6 0 只,在实验环境(室温(21f 2)℃、室内相对湿度(32f 2)%、自然光照)下适应3 d,适应期间饲喂基础饲料,自由进水,第4天早晨停止进食,3 h后称体质量,按体质量分成4 组,每组15 只,单笼饲养,各组间的大鼠平均体质量无显著差异(P>0.05);其中,第1组为基础日粮组(对照组),第2组为市售常温婴儿配方乳I组(简称市售组I),第3组为市售常温婴儿配方乳II组(简称市售组II),第4组为巴氏杀菌婴儿配方乳组(简称实验组)。连续喂养4 周,记录实验开始第0、7、14、21、28天大鼠的体质量。参照GB 10765ü 2010《食品安全国家标准 婴儿配方食品》及GB 10769ü 2010《食品安全国家标准 婴幼儿谷类辅助食品》进行饲料配制[17]。市售组和实验组的基础用料成分无明显差别,各组饲料制作时的配料如表1所示。
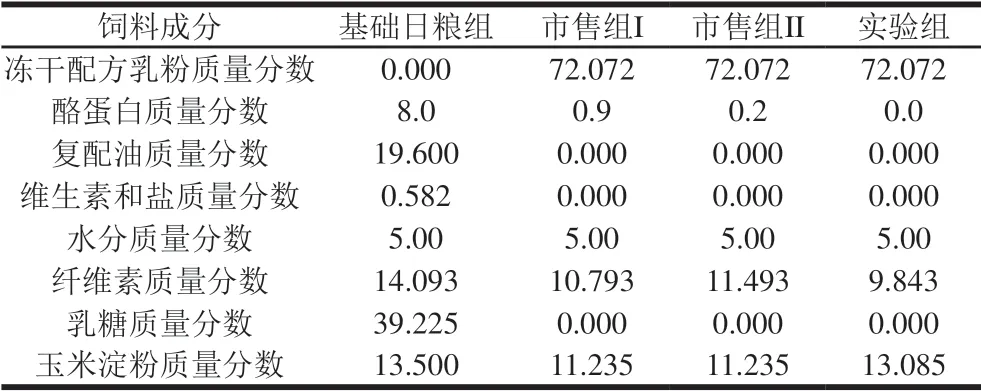
表 1 大鼠饲料的组成Table 1 Rat diet compositions%
1.3.2 大鼠T细胞亚群的测定
于实验第0、28天,每组随机取3 只大鼠,乙醚麻醉后心脏采血,乙二胺四乙酸抗凝,吸取100 μL外周血于EP管中,分别加入1 μL的CD3-FITC、CD4-PE、 CD8a-PerCP单克隆抗体避光染色,反应30 min,每个样品加入体积比1∶10的红细胞裂解液2 mL,混匀,室温静置10 min后,3 000 r/min离心5 min[18]。小心弃去上清液,加入pH 7.4的2 mL PBS,洗涤2 次,弃上清液,加入400 μL PBS混匀,1 h内使用流式细胞仪检测,计算CD3+CD4+T细胞与CD3+CD8+T细胞的比值。
1.3.3 大鼠免疫因子指标的测定
实验开始后,在0、7、14、28 d空腹称体质量,乙醚麻醉后心脏取血5 mL,4 ℃静置12 h,析出血清后,4 ℃、3 000 r/min离心10 min,收集血清分装于EP管中,于-20 ℃保存,根据IL-2、IL-4、IFN-γ、TNF-α、溶菌酶试剂盒说明书步骤测定相应指标水平。
1.4 数据统计与分析
数据均使用Statistix 8软件进行单因素方差分析 (P<0.05表示差异显著),并用Origin 8.5软件作图,结果以平均值±标准差表示。
2 结果与分析
2.1 巴氏杀菌婴儿配方乳对大鼠体质量的影响
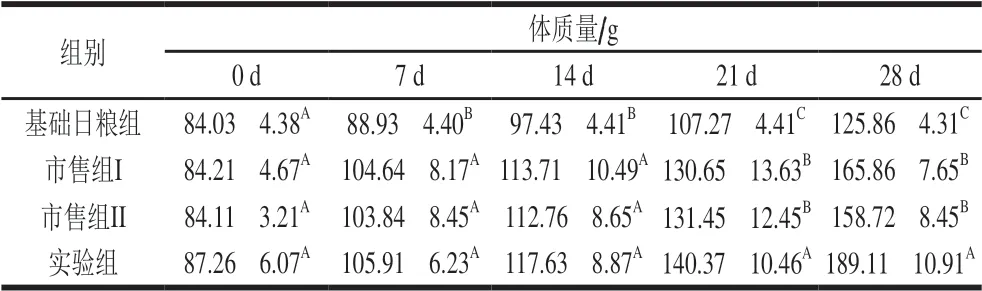
表 2 婴儿配方乳对大鼠体质量变化的影响Table 2 Effect of infant formula on body mass change of rats
由表2可以看出,实验开始前各组大鼠的体质量差异不显著。饲喂两周后,3 组婴儿配方乳组与基础日粮组相比差异显著(P<0.05);3 周后,4 组大鼠的体质量均有不同程度的增长,实验组与对照组和市售组相比差异显著(P<0.05),饲养4 周后,实验组大鼠体质量达到最高,为189.11 g,表明巴氏杀菌婴儿配方乳在促进幼年机体的生长发育方面明显优于市售常温婴儿配方乳。
2.2 巴氏杀菌婴儿配方乳对大鼠血清细胞因子分泌水平的影响
2.2.1 巴氏杀菌婴儿配方乳对大鼠血清IL-2分泌水平的影响

图 1 婴儿配方乳对大鼠IL-2分泌水平的影响Fig. 1 Effect of infant formula on the secretion of IL-2 in rats
IL-2是最早发现的细胞因子之一,又称为T细胞生长因子,对T细胞的增殖、发育、分化、凋亡具有重要的意义[19]。由图1可知,饲喂7 d时,3 组婴儿配方乳组与基础日粮组比差异显著(P<0.05);实验组和市售组II差异显著(P<0.05),与市售组I不显著。随着喂养时间的延长,实验组大鼠血清中IL-2的分泌量逐渐增加,在28 d时达到最大,显著高于其他3 组(P<0.05)。表明实验组与市售组相比,最大程度地保留了免疫物质的生物活性,增加了大鼠血清中IL-2的分泌量,改善大鼠免疫功能,增强机体免疫力。
2.2.2 巴氏杀菌婴儿配方乳对大鼠IL-4分泌水平的影响

图 2 婴儿配方乳对大鼠IL-4分泌水平的影响Fig. 2 Effect of infant formula on the secretion of IL-4 in rats
IL-4主要由活化的T细胞和单核细胞分泌,具有刺激B细胞生长增殖、促进和调节免疫球蛋白E生成、促进T细胞生长增殖、抑制Th1细胞功能等多种生物学功能,对很多炎症反应有促进作用[20]。从图2可以看出,饲养7 d时,实验组和市售组I与基础日粮组相比差异显著 (P<0.05),而这两组之间差异不显著。随着饲养时间的延长,3 组大鼠血清IL-4质量浓度呈降低趋势,原因可能是IL-2分泌量的增加抑制了IL-4的分泌。饲喂14 d时4 组差异不显著;28 d时,市售组I和实验组与基础日粮组相比差异显著(P<0.05),而这两组之间差异不显著,市售组II与实验组差异显著(P<0.05)。结果表明,实验组有降低大鼠血清中IL-4质量浓度的趋势,诱导了以Th1细胞为主导的免疫应答。说明实验组与市售组比能促进免疫细胞对IL的合成和分泌,有效增强大鼠机体免疫力。
2.2.3 巴氏杀菌婴儿配方乳对大鼠IFN-γ分泌水平的影响
IFN-γ是一种由单核细胞和淋巴细胞产生的多功能活性蛋白质[21],主要通过激活单核吞噬细胞来达到杀灭微生物及肿瘤细胞、激活中性粒细胞、强化自然杀伤细胞对肿瘤细胞的杀伤活性、促进巨噬细胞的吞噬功能、抑制宿主向肿瘤细胞提供重要的营养因子或生长因子,从而造成肿瘤细胞的凋亡[22]。因此,本实验通过测定大鼠血清中IFN-γ的分泌水平来对比评估巴氏杀菌婴儿配方乳对大鼠免疫功能的影响,结果如图3所示。
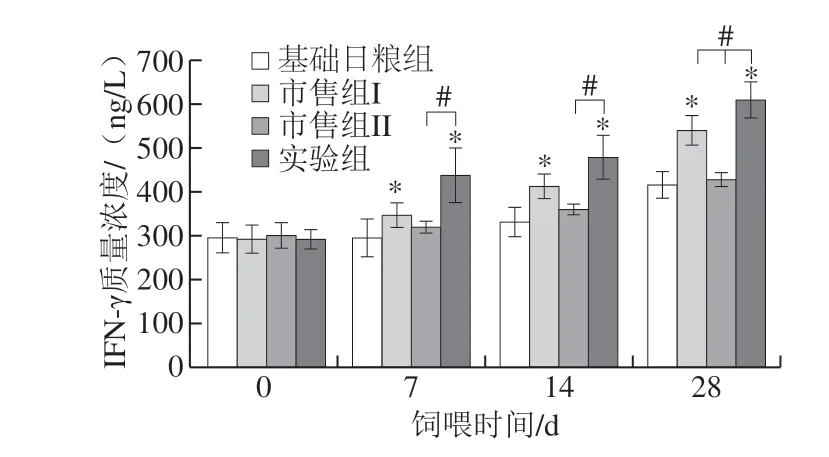
图 3 婴儿配方乳对大鼠IFN-γ分泌水平的影响Fig. 3 Effect of infant formula on the secretion of IFN-γ in rats
由图3可知,在饲喂7 d和14 d时,实验组和市售组I与基础日粮组相比差异显著(P<0.05),市售组II与基础日粮组相比差异不显著;饲喂28 d时,实验组大鼠IFN-γ质量浓度达到最大,显著高于其他3 组 (P<0.05)。随着饲喂时间的延长,IFN-γ质量浓度逐渐增加,原因可能是由于IL-2分泌量的增加促进了IFN-γ的分泌。同时也有研究证明,IFN-γ是细胞介导免疫反应的一个主要标志,具有广泛的免疫调节作用。IFN-γ通过促进抗原特异性使T细胞分化成Th1细胞,进而促进Th1细胞的增殖来增强细胞免疫活性;通过降低Th2细胞活性来发挥抑制体液免疫应答的作用[23-24]。在本研究中,实验组有增加大鼠血清中IFN-γ质量浓度的趋势(P<0.05),说明与市售组相比,实验组极大地保留了免疫物质的活性,提高了大鼠的免疫防御功能。且随着饲喂时间的延长调节作用更加明显。
2.2.4 巴氏杀菌婴儿配方乳对大鼠TNF-α分泌水平的影响
TNF-α主要由巨噬细胞产生,某些T细胞也可分泌,其主要生物活性为直接杀伤肿瘤细胞,亦可促进B细胞的增殖,在许多重要的活性代谢组织如脂肪组织、肝脏和骨骼肌细胞上均有TNF-α的特异性受体[25]。

图 4 婴儿配方乳对大鼠TNF-α分泌水平的影响Fig. 4 Effect of infant formula on the secretion of TNF-α in rats
由图4可知,实验初期4 组TNF-α的分泌水平没有显著性差异;饲喂14 d和28 d时,3 组婴儿配方乳组与基础日粮组相比差异显著(P<0.05)。实验结果说明巴氏杀菌婴儿配方乳与市售常温婴儿配方乳相比能显著提高血清中TNF-α的分泌水平,显著改善大鼠的免疫功能。
综上所述,细胞因子之间具有相互诱生、相互抑制、相互协同和相互拮抗作用。研究证明IL-2可诱导Th1细胞增殖,IL-4则可诱导Th2细胞增殖,两者之间存在一定的相互抑制作用,Th1细胞分泌的IFN-γ可降低IL-4分泌水平[26],IL-2又能够激活T细胞亚群分泌IFN-γ、TNF-α等细胞因子,增强Tc细胞、自然杀伤细胞等的杀伤活性,这与本研究结果一致。巴氏杀菌婴儿配方乳可以使免疫向以Th1细胞为主导的免疫应答偏移[27]。说明巴氏杀菌婴儿配方乳与市售常温婴儿配方乳相比更大地保留了免疫物质的活性,提高了大鼠的免疫防御功能。且随着饲喂时间的延长调节作用更加明显。
2.3 巴氏杀菌婴儿配方乳对大鼠血清溶菌酶分泌水平的影响
血清中的溶菌酶由巨噬细胞、单核细胞和中性粒细胞分泌形成,存在于动物体液及组织中。溶菌酶在机体的修复再生、炎症过程等调节中发挥重要作用,可以从不同角度反映细胞免疫功能和机体抗感染能力[28]。
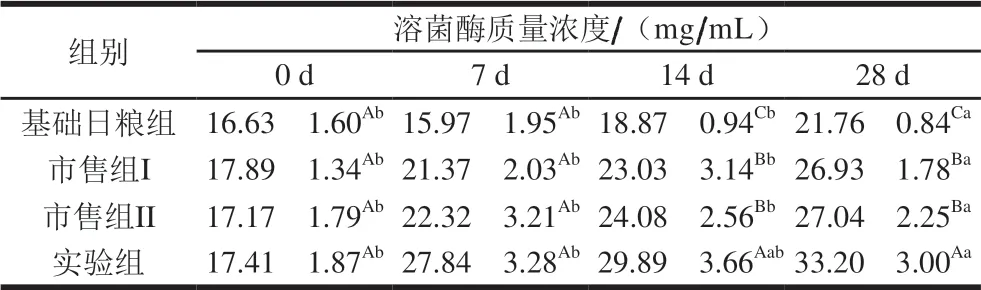
表 3 婴儿配方乳对大鼠血清溶菌酶分泌水平的影响Table 3 Effect of infant formula on serum lysozyme secretion in rats
由表3可知,实验初期,4 组大鼠血清溶菌酶分泌量没有显著差异;随着实验时间延长,饲喂14 d时,与基础日粮组相比,3 组婴儿配方乳组均可以显著提高大鼠血清中溶菌酶质量浓度;饲喂28 d时,实验组大鼠血清溶菌酶质量浓度达到最高,且与市售组差异显著(P<0.05),说明巴氏杀菌婴儿配方乳与市售常温婴儿配方乳相比,可以显著提高大鼠血清中溶菌酶质量浓度,提高机体抗感染的能力,增强机体的非特异性免疫能力,并且随着时间延长效果增强。
2.4 巴氏杀菌婴儿配方乳对大鼠外周血T细胞亚群的影响
T细胞具有不同的功能亚群。CD3是T细胞表面的主要特征性标志物,根据功能和主要组织相容性复合体(major histocompatibility complex,MHC)分子限制性的不同,可将成熟的T细胞分为CD3+CD4+和CD3+CD8+两个细胞亚群[29-30]。CD4是Th细胞和Ti细胞的重要表面标志,为MHC-II类分子的受体,可加强CD4+T细胞与抗原提呈细胞间的黏附力,使T细胞抗原受体与抗原的结合更加稳定。CD8分子主要分布于Tc细胞和Ts细胞,它是MHC-I类分子的受体,能够加强T细胞抗原受体与提呈细胞抗原的结合,促进细胞磷酸化。通过流式细胞仪检测,大鼠外周血淋巴细胞亚群的测定结果见表4。
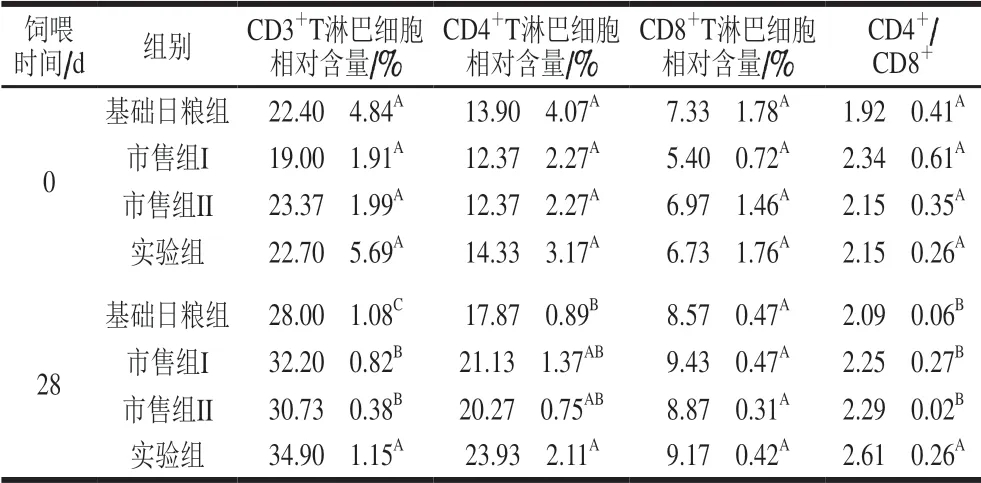
表 4 婴儿配方乳外周血T细胞亚群的影响Table 4 Effect of infant formula on T cell subsets in peripheral blood
通过对4 组数据进行方差分析,结果显示,饲喂28 d时实验组SD大鼠的CD3+T淋巴细胞相对含量与对照组和市售组相比差异显著(P<0.05),表明巴氏杀菌婴儿配方乳可提高SD大鼠的CD3+T淋巴细胞相对含量,可能会使SD大鼠的Ti细胞增多,并对抗原识别、信号转导以及机体快速抗感染免疫起到辅助作用。实验组SD大鼠的CD4+T淋巴细胞相对含量与基础日粮组比差异显著 (P<0.05),表明巴氏杀菌婴儿配方乳可提高SD大鼠的CD4+T淋巴细胞相对含量,对机体免疫调节起一定的辅助作用;而CD8+T淋巴细胞的相对含量不断增加,表明巴氏杀菌婴儿配方乳可能有助于免疫网络的快速形成,并增强机体细胞免疫作用。T细胞作为细胞免疫重要的调节成分,具有上调和下调免疫应答的功能,其免疫应答功能的实现是依靠CD4+、CD8+T淋巴细胞间的相互作用使机体处于免疫平衡状态,所以CD4+/CD8+可作为初步评估机体免疫状态的指标。根据资料显示,正常人外周血中CD4+/CD8+介于0.60~3.45之间[31],4 组SD大鼠血液中CD4+/CD8+均在正常范围内,饲喂28 d时,与对照组和市售组相比,实验组SD大鼠CD4+/CD8+差异显著 (P<0.05),表明其免疫应答的正调节占优势[32]。
3 结 论
巴氏杀菌婴儿配方乳作为一种新型婴幼配方食品填补了国内市场的空白,本实验通过动物喂养实验,以2 款市售常温婴儿配方乳为对照,对比评估了巴氏杀菌婴儿配方乳对大鼠血液免疫因子水平及T细胞亚群的影响。巴氏杀菌婴儿配方乳与市售常温婴儿配方乳比较能够显著提高SD大鼠的体质量,促进大鼠生长发育。巴氏杀菌婴儿配方乳能提高大鼠血清中细胞因子IL-2、IFN-γ、TNF-α及溶菌酶质量浓度,且显著高于市售组及基础日粮组;随着饲喂时间的延长调节作用更加显著。巴氏杀菌婴儿配方乳上调了SD大鼠外周血中CD3+、CD4+、CD8+T淋巴细胞的相对含量以及CD4+/CD8+,且CD4+/CD8+处于正常范围。巴氏杀菌婴儿配方乳与市售常温婴儿配方乳比较,能够促进SD大鼠生长及淋巴细胞的增殖分化,增强体液免疫和细胞免疫功能,并辅助机体免疫调节。
猜你喜欢
工业设计(2022年5期)2022-06-11
农家致富顾问·上半月(2021年6期)2021-12-26
中国果业信息(2021年7期)2021-12-01
中国现代医药杂志(2020年10期)2020-12-14
中国卫生(2016年3期)2016-11-12
中国乳业(2016年4期)2016-11-07
天津科技大学学报(2016年1期)2016-02-28
中国洗涤用品工业(2015年11期)2015-02-28
英语学习(2015年12期)2015-02-01
小说月刊(2014年7期)2014-04-18
