
青稞种质资源表型性状的遗传多样性分析及综合评价
2020-01-14 02:40白羿雄郑雪晴姚有华姚晓华吴昆仑
中国农业科学 2019年23期
白羿雄,郑雪晴,姚有华,姚晓华,吴昆仑
青稞种质资源表型性状的遗传多样性分析及综合评价
白羿雄1,2,郑雪晴1,姚有华1,姚晓华1,吴昆仑1
(1青海大学农林科学院/青海省农林科学院/青海省青稞遗传育种重点实验室/国家麦类改良中心青海青稞分中心,西宁 810016;2西北农林科技大学农学院,陕西杨凌 712100)
【】解析青稞种质资源表型形状的遗传多样性关系及筛选综合性状优异的青稞种质,可为青稞育种及重要性状深入研究提供理论依据。通过对205份青稞15个表型性状采用Shannon-Wiener’s多样性指数进行遗传多样性分析。分析参试青稞种质15个表型性状在西宁试验点和海北试验点的频次分布规律。对各性状利用相关性分析以明确性状间联系;对参试种质进行聚类分析以明确其分类情况。利用主成分分析构建青稞种质综合评价体系,并通过线性回归分析对该体系进行验证。依据综合评价及丰产稳产性分析结果以筛选优异青稞种质。倒伏率遗传变异程度最丰富而重心位置遗传变异最匮乏,海北点遗传变异程度高于西宁点。穗重遗传多样性最丰富,倒伏率遗传单一性最高。除倒伏率外各性状指标均呈正态或偏态分布,分布频次呈中间高两边低的分布趋势;穗长、穗重在各基因型中呈正态分布。各表型性状受环境、基因型、年份影响极显著,且各表型性状的基因型与环境(G×E)、基因型与年份(G×Y)、基因型×环境×年份(G×E×Y)的互作效应均呈极显著。青稞根系、茎秆和穗部组织内各指标间存在显著相关性,且各组织间的农艺性状也存在显著相关性。根系发达、茎部抗折力高的青稞种质其机械固持能力强,倒伏率低;严重倒伏会限制青稞种质穗部生长发育,使穗长变短、穗粒数减少、籽粒变小、穗重变轻、进而使产量锐减。聚类结果表明参试种质可分为三类,第一类是高重心、易倒伏、其余性状居中的种质;第二类是矮秆、低重心且其余性状表现良好的优异种质;第三类是株高较高、根系欠发达、茎部易折、穗部性状表现差的种质。结合值和丰产稳产性分析结果筛选出综合性状表现优异且有较高丰产稳产性的青稞种质5份。参试青稞种质资源的遗传多样性丰富;穗长、穗重在各基因型中呈正态分布,除倒伏率外,其余12个性状在基因型中呈偏态分布;根干重、重心位置、茎秆壁厚、茎粗、茎秆强度、穗长、穗粒数、产量8个指标可作为核心种质评价的综合指标。
青稞;表型性状;遗传多样性;聚类分析;综合评价
0 引言
【研究意义】青稞(L.var.Hook.f.)属禾本科大麦属,因其内外稃与颖果分离,籽粒裸露,故又称裸大麦[1],是栽培大麦变种。青稞为青藏高原最具特色的农作物,具有耐寒、抗旱、抗逆性强、适应性广、生育期短等特点[2]。青稞主要分布在中国五省藏区,其中青海省种植面积达10.67万公顷,占总耕地面积19%左右,青稞在藏区总种植面积高达25.58万公顷[3],是藏族同胞的主粮[4]。青稞因具有较高的蛋白质、高膳食纤维、高β-葡聚糖并含有丰富的多酚类物质而被用于食品加工业[5],青稞秸秆是牧区牦牛和藏羊的主要饲草,对藏区畜牧业发展至关重要[6]。【前人研究进展】遗传多样性研究及综合评价对掌握物种遗传信息具有推动作用,也为育种工作提供便利。近年来,科研工作者主要采用形态标记[7]、生化标记[8]和分子标记[9-11]等手段开展作物遗传多样性研究和种质资源的综合评价。以SSR标记对青稞多样项研究结果表明,青稞种质在农艺性状中表现出丰富的遗传多样性[12-13],不同生态区青稞材料在遗传上存在较大差异[14],野生种质中等位基因数量高于栽培大麦[15]。以SNP标记对种质群体结构及性状的研究结果表明,可将33 176份大麦种质划分为5个亚群,且发现许多参与大麦驯化和育种选择的特征标记[16],二棱和六棱大麦间差异较大[17]。表型性状和分子标记是育种家获取不同青稞资源遗传差异的重要途径,表型性状观察分类是青稞分类学的主要研究方法之一。目前,在水稻[18]、谷子[19]、陆地棉[20]、藜麦[21]、花生[22]中以表型性状开展遗传多样性和综合评价的研究已有相关报道,开展种质资源遗传多样性研究和综合评价对深化种质利用具有重要意义。目前,中国青稞特异性、一致性、稳定性(distinctness uniformity stability,DUS)测试指南以形态性状测试为主,且详细界定了用于品种鉴别的表型及观测时期。青稞为自花授粉作物,新品种培育以传统种间杂交选育为主,利用分子标记辅助选择技术和转基因技术进行育种的单位较少。青稞育种过程因优异亲本数量有限导致其遗传基础狭窄,严重制约优异品种的选育进程。为进一步提高育成青稞品种产量、品质及抗性水平,需收集并筛选优异种质。【本研究切入点】目前,对青稞表型性状在自然群体间的分布规律尚不明确,且适宜用于青稞优异种质资源评价和筛选的指标体系尚未见相关报道。【拟解决的关键问题】本研究拟对两年两点间205份青稞种质的15个表型性状进行研究,并通过相关性分析、主成分分析、聚类分析和逐步线性回归分析明确该种质群体结构,并构建青稞优异种质评价体系,筛选出丰产稳产青稞种质,以期为新品种选育和优异性状的全基因组关联分析(genome-wide association study,GWAS)奠定理论基础。
1 材料与方法
1.1 试验材料
以国内外205份青稞种质为参试材料(电子附表1),其中含西藏种质111份、青海34份、四川20份、甘肃4份、江苏4份、湖南2份、浙江2份、国外种质2份、源于其他各省种质5份,并有21份种质来源不详。
1.2 试验设计
试验于2017—2018年在青海大学农林科学院(青海省西宁市,河谷灌溉农业区,地理位置:36°43′N,101°45′E,海拔2 261 m)和海北州农业科学研究所(青海省海北州海晏县,高寒农牧交错区,地理位置:37°0′N,100°51′E,海拔3 160 m)试验田内进行。西宁试验地土质为沙壤土,土壤耕层有机质为22.49 g·kg-1、全氮含量1.78 g·kg-1、速效磷含量37.48 mg·kg-1、速效钾含量286 mg·kg-1。海北试验地土壤为高山草甸土,土壤耕层有机质为40.73 g·kg-1,全氮含量2.37 g·kg-1、速效磷含量14.50 mg·kg-1、速效钾含量86.13 mg·kg-1。根据生产上青稞品种的施肥量,两点间均施用磷酸二氢铵300 kg·hm-2和尿素150 kg·hm-2,苗期追施尿素37.5 kg·hm-2,等行距株距人工点播,行距20 cm,株距5 cm。2017年,西宁试点于4月6日播种,8月5日收获,海北试点于4月15日播种,9月20日收获;2018年,西宁试点于4月8日播种,8月10日收获,海北试点于4月17日播种,9月23日收获。试验采用随机区组设计,小区面积为2.5 m×4 m,设3次重复。
1.3 试验方法
每份参试材料选取长势一致的10株材料进行表型性状调查。田间调查青稞倒伏率、产量,根系所测性状为根数和根干重;茎部所测性状包括株高、有效分蘖数、重心位置、茎秆壁厚、茎粗、茎秆强度;穗部所测性状为穗长、穗重、小穗数、穗粒数、千粒重,共统计15个指标。参照白羿雄等[23]研究方法测定表型性状。
1.4 数据分析
采用Excel2010进行遗传多样性分析;采用作物品种区域试验管理分析系统1.0(RCT99)的AMMI模型对参试种质进行丰产稳产性分析[24-25];采用SPSS20.0进行正态性检验、互作分析、相关性分析、聚类分析、主成分分析和逐步线性回归分析,以构建青稞种质资源的表型评价体系。其中聚类分析方法为系统聚类平均联结(组间)法,隶属函数和遗传多样性指数的计算方式如下:
()=(‒min)/(max‒min)(=1,2,3,...,n)
式中,()为各材料第个性状的隶属函数值,为各材料第个性状值,max、min分别为所有参试材料中第个性状的最大值和最小值[26]。
采用Shannon-Wiener’s多样性指数(Shannon- Wiener’s diversity index H′)进行遗传多样性评价。
=–∑×ln(=1,2,3,...,n)
式中,表示某性状第级别内材料份数占总份数的百分比[26]。
2 结果
2.1 表型性状的遗传多样性分析
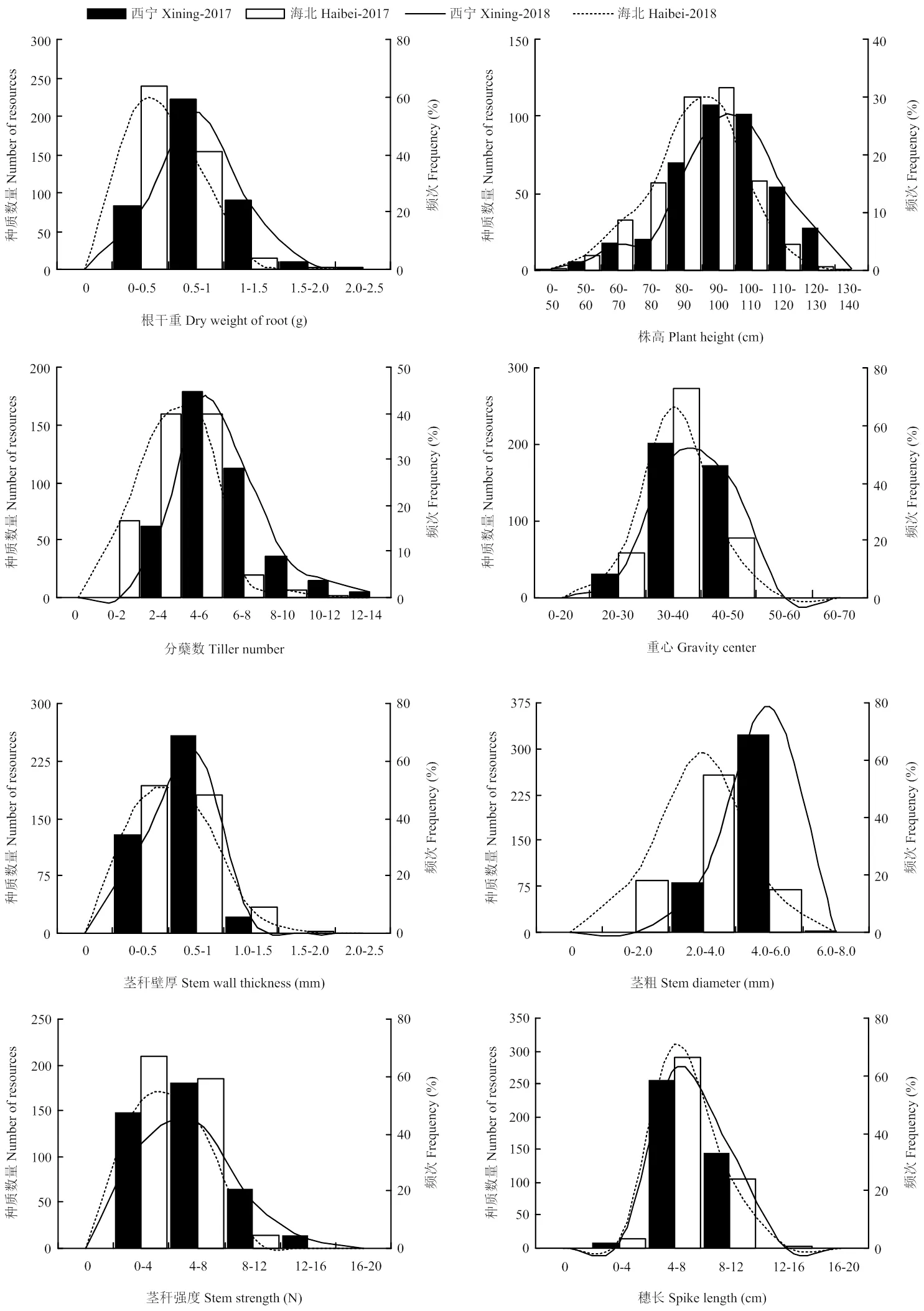
表型性状中倒伏率变异系数最大(=121.49%),表明倒伏率在表型性状中遗传变异最为丰富;重心位置变异系数最小(=14.68%),其遗传变异程度最低。在各性状中穗重的多样性指数最大(=2.08),其遗传多样性最为丰富。多样性指数分布范围为1.41—2.08(表1),且各性状基因型均分布在较大变幅区段内(图1),表明参试种质表型性状的遗传多样性丰富。
各性状于两点间差异较大(表2)。西宁点除千粒重外各参试指标均值高于海北点,而西宁点大部分性状的变异系数低于海北点;表明相较于西宁点,海北点遗传变异更丰富,西宁点更适宜于各基因型生长发育。

表1 青稞种质资源的表型性状
NOR:根数Number of roots;RDW:根干重Root dry weight;PH:株高Plant height;NOT:分蘖数Number of tillers;COG:重心位置Centre-of-gravity position;WT:茎秆壁厚Stem wall thickness;DMS:茎粗Diameter of main stem;SS:茎秆强度Stem strength;SL:穗长Spike length;PW:穗重Panicle weight;SN:小穗数Spikelet number;KPS:穗粒数Kernels per spike;TGW:千粒重Thousand grain weight;Y:产量Yield;LR:倒伏率Lodging rate。下同the same as below

表2 表型性状在不同环境下的变异
2.2 表型性状在各基因型中的分布规律
除倒伏率外各性状指标均呈正态或偏态分布,分布频次呈中间高两边低的分布趋势(图1)。正态检验分析结果表明穗长、穗重在各基因型中呈正态分布;而穗粒数和千粒重基因型在海北点间呈正态分布(表3)。根数、株高、穗长、穗重、穗粒数、倒伏率的最高频次分布区间在两年两点间相同。部分性状最高分布频次在两点间存在一定差异,表明环境因素对各性状分布频次会产生一定程度影响(图1)。
2.3 基因型、年份与环境的互作效应分析
各表型性状受环境因素、基因型、年份的影响均极显著,且各表型性状的基因型与环境(G×E)、基因型与年份(G×Y)、基因型×环境×年份(G×E×Y)间存在极显著互作效应(表4)。
2.4 性状间相关分析
青稞根系、茎秆和穗部组织内各指标间存在显著相关性(表5)。根系中根数和根干重呈极显著正相关性。茎秆中重心位置、茎秆壁厚、茎粗两两指标间均呈极显著正相关性;穗部穗长、穗重、小穗数两两间均呈极显著正相关性,而穗粒数与千粒重间呈极显著负相关性。
表3 参试材料正态性检验
+:表明该性状呈正态分布
+: Indicates the trait showed normally distributed
青稞根系、茎秆和穗部组织间的农艺性状也存在显著相关性。根系两项指标与茎秆中除株高外的各项指标呈极显著相关性,而同穗部除千粒重外各指标呈极显著相关性。茎秆各项指标同穗重呈显著相关性,且部分指标同穗部相关指标呈极显著相关性。产量同除株高性状外的各指标呈极显著正相关性,表明根系发达,茎秆分蘖多抗折力强,穗粒数多、千粒重大的参试材料产量高。倒伏率同根数、根干重、茎秆壁厚、茎秆强度、穗长、穗重、小穗数、穗粒数、千粒重呈极显著负相关性,同产量呈显著负相关性。根系发达、茎部抗折力高的青稞植株其机械固持能力较强,倒伏率低;青稞植株严重倒伏会限制穗部生长发育,使穗长变短、穗粒数减少、籽粒变小、穗重变轻、进而使产量锐减。
2.5 主成分分析及综合评价
前7个主成分代表青稞种质资源表型性状85.02%的遗传信息量(表6)。第1主成分贡献率为31.91%,为穗重、穗粒数因子;第2主成分贡献率为15.58%,为株高因子;第3主成分贡献率为11.69%,为分蘖数因子;第4、5主成分贡献率分别为8.79%和6.30%,均为千粒重因子;第6主成分贡献率为5.83%,为茎粗因子;第7主成分贡献率为4.93%,为穗长因子。
将种质资源的15个表型性状的值利用模糊隶属函数进行标准化处理,并将其带入7个主成分的得分中,求得各种质的7个主成分得分,第1主成分得分是:1=0.6041+0.6642+0.0623+0.2084+0.2755+ 0.5836+0.6077+0.7508+0.6269+0.75810+0.69111+0.75312-0.00813+0.71014-0.26715,利用7个主成分贡献率权重(0.375、0.183、0.138、0.103、0.074、0.069和0.058),构建用于筛选青稞优异种质的评价指标值,其为=0.3751+0.1832+0.1383+0.1034+ 0.0745+0.0696+0.0587,值越高,可初步表明该种质综合性状越好。
利用205份青稞种质在两年两点间值的平均值大小,筛选出排名前十的种质为170(1.498)、131(1.495)、168(1.482)、165(1.464)、118(1.460)、164(1.453)、109(1.446)、143(1.443)、132(1.416)和107(1.415),其值较高,表明其综合性状较好;排名较低的10份种质为116(0.878)、96(0.873)、179(0.873)、124(0.850)、193(0.842)、130(0.819)、6(0.760)、83(0.714)、4(0.688)和5(0.675),其值较低。
以参试材料表型性状及值进行逐步回归分析,并构建线性回归方程以用于优异种质鉴选,构建方程为:=(-344.805+125.1722+12.6205+121.1326+51.4047+18.7368+37.7339+4.14812+0.02714)×10-3,其中2、5、6、7、8、9、12和14分别代表根干重、重心位置、茎秆壁厚、茎粗、茎秆强度、穗长、穗粒数、产量。该回归方程的相关系数=0.988,决定系数2=0.976,表明2、5、6、7、8、9、12、14这8个自变量可决定值总变异的97.6%,值为4 062.225。相关性分析研究结果表明,以上8个表型性状同值均呈极显著相关性(表7),则该方程可用于青稞种质资源的综合评价。
2.6 表型性状的聚类分析
可将205份供试材料划分为3个亚类(图2)。第一类,共177份,占供试材料的86.34%,为高重心、易倒伏、而其它性状居中的种质;第二类,共21份,为矮秆、低重心且其余性状表现良好的优异种质,其中,产量均值高达6 250.65 kg·hm-2;第三类,共7份,为株高较高,而根系欠发达、茎部易折、穗部性状表现较差的种质。

表4 基因型、环境和年份的互作效益分析

变异来源Variance sourcesdf穗长SL穗重PW小穗数SN穗粒数KSP千粒重TGW产量Y倒伏率LR FPFPFPFPFPFPFP 基因型Genotype(G)20410.740.0019.450.0010.330.0025.940.0073.940.003.830.001814.240.00 年份Year(Y) 154.620.0033.260.0041.020.00397.650.001529.210.00961.550.0060624.730.00 环境Environment(E) 1766.820.00451.230.00251.610.001018.160.00828.190.0015.510.0049011.260.00 基因型×环境G×E2042.550.004.040.004.420.004.220.0033.030.002.030.00690.300.00 基因型×年份G×Y2043.360.004.880.003.660.004.500.0030.530.002.290.00510.090.00 基因型×环境×年份G×E×Y2043.340.006.600.005.900.004.010.0039.470.005.480.001450.580.00

表5 青稞15个表型性状相关系数
*,**分别表示在0.05和0.01水平上显著相关。下同 *, ** indicate significant correlation at the 0.01 and 0.05 levels, respectively. The same as below

表6 青稞种质资源表型性状的主成分分析
E:特征值Eigenvalue;CR:贡献率Contribution rate;CCR:累计贡献率cumulative contribution rate

图2 205份青稞种质聚类分析图

表7 表型性状与综合得分(F值)间的相关性
2.7 丰产稳产性分析
从图3可以看出,参试种质中ZDM9845的丰产性最好,但其稳产性较差;ZYM1906和ZYM1757的稳产性较好,但丰产性极差。甘青4号则具有最好的丰产稳产性,另外昆仑14号、ZDM9734、ZDM9334和西藏42的丰产稳产性均较好。
丰产稳产性较好的5个品种为甘青4号、昆仑14号、藏815007、ZDM9334和西藏42,其值在参试材料中位于前十且均属于聚类分析中优异青稞种质(第二类)。聚类分析中优质青稞种质类别(第二类)中包含值最高的10份品种中的9份,故聚类分析结果和AMMI模型共同验证本研究所构建回归方程有较高可靠性。

图3 205份青稞种质的AMMI双标图
3 讨论
3.1 表型性状的遗传多样性分析
表型性状是结构基因的功能表现,是长期自然与人工选择的结果,虽易受时间、环境因素影响,但表型性状观察结果直观,经济实用,通过表型性状的遗传多样性分析已经在多种作物中得到应用[27-28]。本研究利用青稞的15个表型形状,计算Shannon-Wiener’s多样性指数的均值为1.94,遗传多样性指数较高,说明该批种质具有较高的丰富度和均匀度。倒伏率在15个表型性状中变异系数最高,该结果同白羿雄等[23]研究结果一致,说明倒伏率的遗传变异极为较丰富,倒伏率的分布频次中无倒伏和严重倒伏的青稞品种较多,中度倒伏品种较少,歧化现象严重,同时倒伏率在基因型间的互作、环境间的互作均为极显著,表明基因型和环境均会对倒伏率性状的遗传效应产生极大影响,该结果同黄文辉等[29]研究结果一致。本研究结果表明青稞穗长、穗重2个性状在基因型中呈正态分布,同王贵学等[30]在小麦中对这两个性状的研究结果相同。本研究过程受到群体结构、样本数量、环境因素的限制,检验出青稞中2个性状呈正态分布,其余性状是否呈正态分布,仍需开展进一步的研究。
性状指标在西宁和海北两生态区呈现一定差异性,例如东都红胶泥、ZDM9854、甘青9188-10在西宁点表现较好丰产性,适宜在河谷灌溉农业区种植,却不适宜在高寒农牧交错区种植。除千粒重外西宁点各性状指标均高于海北点,该现象可能为环境因子所致。相较于西宁点,海北点海拔高、风力大(海北点平均风速是西宁点的2.3倍)、气温低(图4),会使青稞植株发育受限,最终致使绝大部分指标降低;但高海拔地区昼夜温差大,光合作用累积的营养物质于夜间消耗较少,籽粒中相对积累较多的营养物质,致使海北点青稞千粒重较大,该千粒重的研究结果同侯维海等[31]研究结果一致。

图4 2017—2018西宁、海北气象数据
3.2 青稞种质资源表型性状综合评价
种质资源综合评价可为作物品种选育及综合利用提供重要参考。在作物中常利用加权隶属函数法进行耐盐[32]、耐低磷等耐性及抗性[33]进行综合评价;灰色关联分析、加权分析进行综合评价的方法不能有效评价种质具有一定局限性;主成分分析法结合丰产稳产性分析是近年来对作物开展遗传多样性评价的有效方法[34]。主成分分析基于表型数据标准化,且利用其得分归一化处理的值可为育种和遗传研究提供参考[19]。而本文利用主成分分析和逐步线性回归构建青稞综合评价体系,从15个表型性状中筛选出根数等8个性状作为鉴定指标。以评价体系为基础,结合丰产稳产性分析结果,筛选出5份优异种质,分别为昆仑14号、甘青4号、藏815007、ZDM9334、西藏42号。筛选种质均表现出根系发达,茎秆粗壮,穗部饱满,每穗粒数均高于60,产量高于6 000 kg·hm-2,不倒伏,表型性状综合表现优异、且表现出较好丰产和稳产特性。其中昆仑14号为国审青稞品种,具有高产、稳产、适应性广的特点,其在五省藏区推广面积达13.35万公顷;甘青4号为甘南藏族自治州的主栽品种,其在该州推广面积达5万公顷。由此表明,利用表型综合性状综合得分(值)对青稞材料进行综合评价具有较高的可靠性,且以此方法可增加筛选优良种质的准确率,提高育种工作效率。
此外,表型性状研究会存在一定局限性,为拓宽研究的广度和深度仍需结合现代分子技术手段进行遗传学评价和鉴定[35-37]。笔者所在青稞课题组拟后续结合SNP标记的基因分型结果,进一步明确该群体的遗传结构和亲缘关系远近,以拓展青稞遗传多样性和综合评价研究的深度和广度。
4 结论
参试青稞种质资源的遗传多样性丰富,穗长、穗重在各基因型中呈正态分布,其余12个性状在基因型中呈偏态分布;该批种质可划分为3个亚类;根干重、重心位置、茎秆壁厚、茎粗、茎秆强度、穗长、穗粒数、产量8个指标可作为核心种质评价的综合指标。
[1] 矫晓丽, 迟晓峰, 董琦, 肖远灿, 胡风祖. 青海地区不同品种青稞中B族维生素含量分布. 氨基酸和生物资源, 2011, 33(2): 13-16.
Jiao X L, Chi X F, Dong Q, Xiao Y C, Hu F Z. HPLC determination of vitamin B in different highland barleys from Qinghai province., 2011, 33(2): 13-16. (in chinese)
[2] 夏腾飞, 王蕾, 徐金青, 王寒冬, 张怀刚, 刘登才, 沈裕虎, 昌西. 267份青藏高原青稞种质材料的表型多样性分析. 西北农业学报, 2018, 27(2): 182-193.
Xia T F, Wang L, Xu J Q, Wang H D, Zhang H G, Liu D C, Shen Y H, Chang X. The genotypic diversity analysis of 267 six-rowed hulless barley accessions from the Qinghai-Tibetan plateau., 2018, 27(2): 182-193. (in chinese)
[3] 吕远平, 熊茉君, 贾利蓉. 青稞特性及在食品中的应用. 食品科学, 2005, 26(7): 245-249.
Lü Y P, Xiong M J, Jia L R. Characteristics of barley and application in food industry., 2005, 26(7): 245-249. (in chinese)
[4] 刘三才. 我国青稞的质量与品质研究进展. 作物杂志, 2014(4): 1-5.
Liu S C. Progress in quality of naked barley in China., 2014(4): 1-5. (in chinese)
[5] 刘小娇, 王姗姗, 白婷, 靳玉龙, 文华英, 张玉红. 青稞营养及其制品研究进展. 粮食与食品工业, 2019(1): 43-47.
Liu X J, Wang s s, Bai T, JIN Y L, Wen H Y, Zhang Y H. Advance of hulless barley nutrition and its products., 2019(1): 43-47. (in chinese)
[6] 李龙兴. 西藏主要农作物秸秆与牧草混合青贮的研究[D]. 南京: 南京农业大学, 2013.
Li L X. Study on the mixed silage of main crops straw and forage in Tibet[D]. Nanjing: Nanjing Agricultural University, 2013. (in chinese)
[7] Zanklan A S, Becker H C, Sorensen M, Pawelzik E, Grüneberg W J. Genetic diversity in cultivated yam bean (spp.) evaluated through multivariate analysis of morphological and agronomic traits., 2018, 65(3): 811-843.
[8] Yin Y Q, Ma D Q, Ding Y. Analysis of genetic diversity of hordein in wild close relatives of barley from Tibet., 2003, 107(5): 837-842.
[9] Würschum T, Langer S M, Longin C F H, Korzun V, Akhunov E, Ebmeyer E, Schachschneider R, Schacht J, Kazman E, Reif J C. Population structure, genetic diversity and linkage disequilibrium in elite winter wheat assessed with SNP and SSR markers., 2013, 126(6): 1477-1486.
[10] Sorkheh K, Masaeli M, Chaleshtori M H, Adugna A, Ercisli S. AFLP-based analysis of genetic diversity, population structure, and relationships with agronomic traits in rice germplasm from north region of IRAN and world core germplasm set., 2016, 54(2): 177-193.
[11] Zhou R, Wu Z, Cao X, Jiang F L. Genetic diversity of cultivated and wild tomatoes revealed by morphological traits and SSR markers., 2015, 14(4): 13868-13879.
[12] 胥婷婷. 青稞种质资源遗传多样性分析和核心种质的构建[D]. 杭州: 浙江师范大学, 2012.
Xu T T. The genetic diversity analysis and establishment of core collection in hulless barley germplasm resources[D]. Hangzhou: Zhejiang Normal University, 2012. (in chinese)
[13] 孟亚雄, 孟祎林, 汪军成. 青稞遗传多样性及其农艺性状与SSR标记的关联分析. 作物学报, 2016, 42(2): 26-35.
Meng Y X, Meng Y L, Wang J C. Genetic diversity and association analysis of agronomic characteristics with SSR markers in hulless barley., 2016, 42(2): 26-35. (in chinese)
[14] 吴昆仑. 青稞种质资源的SSR标记遗传多样性分析. 麦类作物学报, 2011, 31(6): 1030-1034.
Wu K L. Genetic diversity analysis of hulless barley germplasm by SSR marker., 2011, 31(6): 1030-1034. (in chinese)
[15] Shakhatreh Y, Baum M, Haddad N, ALRAABAH M, CECCARELLI S. Assessment of genetic diversity among Jordanian wild barley () genotypes revealed by SSR markers., 2016, 63(5): 813-822.
[16] Muñoz-Amatriaín M, Cuesta-Marcos A, Endelman J B, Alrababah M, Ceccarelli S. The USDA barley core collection: genetic diversity, population structure and potential for genome-wide association studies., 2014, 9(4): e94688.
[17] Amezrou R, Gyawali S, Belqadi L, Chao S M, Arbaoui M, Mamidi S, Rehman S, Sreedasyam A, Verma P S. Molecular and phenotypic diversity of ICARDA spring barley (L.) collection., 2018, 65(1): 255-269.
[18] 胡标林, 万勇, 李霞, 雷建国, 罗向东, 严文贵, 谢建坤. 水稻核心种质表型性状遗传多样性分析及综合评价. 作物学报, 2012, 38(5): 829-839.
Hu B L, Wan Y, Li X, Lei J G, Luo X D, Yan W G, Xie J K. Analysis on genetic diversity of phenotypic traits in rice () core collection and its comprehensive assessment., 2012, 38(5): 829-839. (in chinese)
[19] 王海岗, 贾冠清, 智慧, 温琪汾, 董俊丽, 陈凌, 王君杰, 曹晓宁, 刘思辰, 王纶, 乔治军, 刁现民.谷子核心种质表型遗传多样性分析及综合评价.作物学报,2016, 42(1):19-30.
Wang H G, Jia G Q, Zhi H, Wen Q F, Dong J L, Chen L, Wang J J, Cao X N, Liu S C, Wang L, Qiao Z J, Diao X M. Phenotypic diversity evaluations of foxtail millet core collections., 2016, 42(1): 19-30. (in chinese)
[20] 代攀虹, 孙君灵, 何守朴, 王立如, 贾银华, 潘兆娥, 庞保印, 杜雄明, 王谧. 陆地棉核心种质表型性状遗传多样性分析及综合评价. 中国农业科学, 2016, 49(19): 3694-3708.
Dai P H, Sun J L, He S P, Wang L R, Jia Y H, Pan Z E, Pang B Y, Du X M, Wang M. Comprehensive evaluation and genetic diversity analysis of phenotypic traits of core collection in upland cotton., 2016, 49(19): 3694-3708. (in chinese)
[21] 王艳青, 李春花, 卢文洁, 孙道旺, 尹桂芳, 陆平, 王莉花. 135份国外藜麦种质主要农艺性状的遗传多样性分析. 植物遗传资源学报, 2018, 19(5): 1-12.
Wang Y Q, Li C H, Lu W J, Sun D W, Yin G F, Lu P, Wang L H. Genetic diversity analysis of major agronomic traits in 135 foreign Quinoa germplasm accessions., 2018, 19(5): 1-12. (in chinese)
[22] 孙东雷, 卞能飞, 陈志德, 邢兴华, 徐泽俊, 齐玉军, 王幸, 王晓军, 王伟. 花生种质资源表型性状的综合评价及指标筛选. 植物遗传资源学报, 2018, 19(5): 865-874.
Sun D L, Bian N F, Chen Z D, Xing X H, Xu Z J, Qi Y J, Wang X, Wang X J, Wang W. Comprehensive evaluation and index screening of phenotypic traits in peanut germplasm resources., 2018, 19(5): 865-874. (in chinese)
[23] 白羿雄, 姚晓华, 姚有华, 吴昆仑. 青稞抗倒伏性状的基因型差异. 中国农业科学, 2019, 52(2): 228-238.
Bai Y X, Yao X H, Yao Y H, Wu K L.Difference of traits relating to lodging resistance in hulless barley genotypes., 2019, 52(2): 228-238. (in chinese)
[24] 王志龙, 程加省, 杨金华, 乔祥梅, 王志伟, 程耿, 于亚雄. 环境与基因型及其互作对云南早秋大麦产量的影响. 农业开发与装备, 2019(2): 92-93.
Wang Z L, Cheng J S, Yang J H, Qiao X M, Wang Z W, Cheng G, Yu Y X. Effects of environment, genotypes and their interaction on the yield of early autumn barley in Yunnan province., 2019(2): 92-93. (in chinese)
[25] 吴显, 马俊, 梁黔云, 先新, 孙开利, 李世吉, 李喆, 王习秀. AMMI双坐标分析在玉米品种比较试验中的应用. 现代农业科技, 2018(20): 24, 27.
Wu X, Ma J, Liang Q Y, Xian X, Sun K L, Li S J, Li J, Wang X X. Application of AMMI double coordinate analysis in maize varieties comparison test., 2018(20): 24, 27. (in chinese)
[26] Strong W L. Biased richness and evenness relationships within Shannon–Wiener index values., 2016, 67: 703-713.
[27] 余斌, 杨宏羽, 王丽. 引进马铃薯种质资源在干旱半干旱区的表型性状遗传多样性分析及综合评价. 作物学报, 2018, 44(1): 63-74.
Yu B, Yang H Y, Wang L. Genetic diversity analysis and comprehensive assessment of phenotypic traits introduced potato germplasm resources in arid and semi-arid area., 2018, 44(1): 63-74. (in chinese)
[28] 董承光, 王娟, 周小凤, 马晓梅, 李生秀, 余渝, 李保成. 基于表型性状的陆地棉种质资源遗传多样性分析. 植物遗传资源学报, 2016, 17(3): 438-446.
Dong C G, Wang J, Zhou X F, Ma X M, Li S X, Yu Y, Li B C. Evalution on genetic diversity of cotton germplasm resources (L.on morphological characters., 2016, 17(3): 438-446. (in chinese)
[29] 黄文辉, 王会, 梅德圣. 农作物抗倒性研究进展. 作物杂志, 2018(4): 13-19.
Huang W H, Wang H, Mei D S. Research progress on lodging resistance of crops., 2018(4): 13-19. (in chinese)
[30] 王贵学, 邱厥. 从小麦数量性状的表型值分布看正态性检验的必要性——几种正态性检验方法的比较. 西南农业学报, 1990(2): 33-38.
Wang G X, Qiu J. On the necessity of normality test based on the distribution of phenotypic values for quantitative characters of wheat-a comparison of several kinds of normality test., 1990(2): 33-38. (in chinese)
[31] 侯维海, 王建林, 旦巴, 胡单. 不同生态因子条件下西藏青稞种子表型性状的相关分析. 核农学报, 2017, 31(10): 2063-2071.
Hou W H, Wang J L, Dan B, Hu D. Phenotypic correlation analysis of hulless barley kernel traits from Tibetan Plateau region under the condition of different ecological environment., 2017, 31(10): 2063-2071(in chinese)
[32] 陈新, 张宗文, 吴斌. 裸燕麦萌发期耐盐性综合评价与耐盐种质筛选. 中国农业科学, 2014, 47(10): 2038-2046.
Chen X, Zhang Z W, Wu B. Comprehensive evaluation of salt tolerance and screening of salt tolerant germplasm in naked oats during germination., 2014, 47(10): 2038-2046. (in chinese)
[33] 张建华, 金黎平, 谢开云, 庞万福, 卞春松, 段绍光, 屈冬玉. 不同基因型马铃薯块茎损伤性状的综合评价. 中国农业科学, 2009, 42(1): 198-203.
Zhang J H, Jin L P, Xie K Y, Pang W F, Bian C S, Duan S G, QU D Y. Comprehensive evaluation of tuber damage traits in different genotypes of potato., 2009, 42(1): 198-203. (in chinese)
[34] 陶爱芬, 祁建民, 林培青, 方平平, 吴建梅, 林荔辉. 红麻优异种质产量和品质性状主成分聚类分析与综合评价. 中国农业科学, 2008(9): 2859-2867.
TAO A F, Qi J M, LIN P Q, FANG P P, WU J M, LIN L H. Cluster analysis and evaluation of elite kanaf germplasm based on principal components., 2008(9): 2859-2867. (in chinese)
[35] Li C Q, Song L, Zhu Y J, Zhai Y J, Wang Q L. Genetic diversity assessment of upland cotton variety resources in china based on phenotype traits and molecular markers., 2017, 57(1): 290-301.
[36] Li Y, Colleoni C, Zhang J, Liang Q, Hu Y, Ruess H, Schmitt E. Genomic analyses yield markers for identifying agronomically important genes in potato., 2018, 11(3): 473-484.
[37] Boczkowska M, Zebrowski J, Nowosielski J, Kordulasińska I, Nowosielska D, Podyma W. Environmentally related genotypic, phenotypic and metabolic diversity of oat (L.) landraces based on 67 Polish accessions., 64(8): 1829-1840.
Genetic Diversity Analysis and Comprehensive Evaluation of Phenotypic Traits in Hulless Barley Germplasm Resources
BAI Yixiong1,2, ZHENG Xueqing1, YAO Youhua1, YAO Xiaohua1, WU Kunlun1
(1Academy of Agriculture and Forestry of Qinghai University/Qinghai Academy of Agriculture and Forestry sciences/Qinghai Provincial Key Laboratory of Hulless Barley Genetics and Breeding/Qinghai Hulless Barley Sub-center of National Triticeae Improvement Center, Xining 810016;2College of Agronomy, Northwest A & F University, Yangling 712100, Shaanxi)
【】The genetic diversity in the hulless barley (L. var.Hook. f.) germplasm resources can be screened to improve phenotypic appearance traits. 【】Shannon-Wiener diversity index was used to analyze thegenetic diversity of 15 phenotypic traits in 205 hulless barley accessions. The distribution frequency of all phenotypic traits in test germplasm resources were analyzed, which contained the data of Xining and Haibei experiment point. Correlation analysis was used to identify the relationship among all traits; cluster analysis was carried out to clarify its classification of the tested germplasm. Principal component analysis was used to construct a comprehensive evaluation system of hulless barley germplasm resources, and the system was verified by linear regression analysis. Screening excellent hulless barley germplasm resources based on the results of comprehensive evaluation and high yield and stability analysis. Correlation analysis, principal component analysis, cluster analysis and high yield and stability analysis were used to evaluate the germplasms. 【】The genetic variation level of lodging rate was the richest, the genetic variation level of the center of gravity is the most deficient, the genetic variation of the Haibei test plot showed higher than that of Xining point. The genetic diversity of panicle weight was the most abundant, the genetic uniformity of lodging rate was the highest. Except for the lodging rate, the additional traits showed normal or skewed distribution, and the distribution frequency showed the trend of higher middle and lower sides. The spike length and spike weight showed normally distributed in each genotype. Spike length and panicle weight have normal distributions among the hulless barley accessions. The phenotypic traits showed highly significant differences related to environmental, genotypes, and years factors. Genotype and environment (G×E) factors, genotype and year (G×Y) factors and genotype × environment× year (G×E×Y) interactions produced highly significant differences in the phenotypic traits. There was a significant correlation between the indicators among the roots, stems and panicles of the hulless barley, and there was also a significant correlation between the agronomic traits among the tissues. Well-developed root system, greater stem bending resistance, and stronger mechanical retention, reduced the lodging. Susceptibility to lodging of hulless barley limits the growth and development of spikes and reduce spike length, grain number per ear, grain size, and ear weight. This greatly reduces hulless barley yield. The clustering results demonstrated that the germplasm could be divided into three categories. Category one had a high center of gravity and was susceptible to lodging, but the remaining traits were intermediate. Category two contained germplasm with good agronomic qualities such as dwarf, stalk with a low center of gravity and other useful traits. Category three contained germplasm with high plant height, underdeveloped roots, easily to folded stems, and poor panicle performance. The F value results and yield stability analysis identified five types of barley hull germplasms with excellent overall traits, high yield and stability. 【】 High genetic diversity exists in the barley germplasm resources. The spike length and spike weight were normally distributed within each genotype. With the exception of the lodging rate, 12 other traits had skewed distributed within the genotypes. Eight traits including root dry weight, center of gravity, stem wall thickness, main stem diameter, stem strength, spike length, kernels per spike and yield are useful indicators for evaluating barley germplasms.
hulless barley; phenotypic traits; genetic diversity; cluster analysis; comprehensive evaluation

10.3864/j.issn.0578-1752.2019.23.002
2019-05-28;
2019-07-24
青海省农林科学院创新基金重点研发项目(2019-NKY-01)、国家现代农业产业技术体系(CAS-05)、青海大学中青年科研基金项目(2017-QNY-2)、西藏自治区重大科技专项(XZ201801NA01-014)
白羿雄,E-mail:yixiongbai@163.com。
吴昆仑,E-mail:wklqaaf@163.com
(责任编辑 李莉)
猜你喜欢
麦类作物学报(2022年7期)2022-08-09
江西农业大学学报(2022年3期)2022-07-07
星星·诗歌原创(2022年1期)2022-04-16
中国生态农业学报(中英文)(2021年9期)2021-09-10
农业机械学报(2020年11期)2020-11-25
西藏人文地理(2020年4期)2020-11-19
翠苑(2019年4期)2019-11-11
农村百事通(2018年16期)2018-09-29
江苏农业科学(2016年4期)2016-06-14
安徽农学通报(2015年15期)2015-08-19
