
水分胁迫对梨枣光合特性的影响
2019-12-31 05:15朱惠玲贺玉玉冯晓东
延安大学学报(自然科学版) 2019年4期
朱惠玲,李 丹,贺玉玉,冯晓东,2*
(1.延安大学 生命科学学院;2.陕西省红枣重点实验室,陕西 延安 716000)
在绿色植物生长发育的过程中,水分是不可忽视的影响因子之一,水分过多或者水分亏损都会影响植物的正常生长发育[1]。植物不断地从外界环境中吸收水分,保持体内正常的含水量,使植物能够正常的进行各项生理活动。因此,植物体的正常生命活动过程需要水分的参与,植物就是在不断地吸收、运输和利用水分的过程中维持其正常的生命活动。水分作为生物体的重要组成的部分,如果出现水分亏缺的现象,植物的正常生命活动过程就会受到阻碍,出现萎蔫甚至死亡现象。而且在植物的生长过程中干旱对于植物的伤害是最大的[2],必须保持一定的含水量才有利于植物的生长。
梨枣(ZiziphusjujubemillLiZao)为鼠李科(Rhamnaceae)枣属(ZizyphusMill)植物,原产于我国,是我国特有的经济树种,近年来已广泛种植,为枣树中稀有的名贵鲜食品种,具有耐旱、喜光、适应性强等特点[3]。本试验以3年生梨枣为材料,初步研究水分胁迫对3年生梨枣光响应特征的影响,旨在了解植物在水分胁迫下对光的响应机理,比较分析水分胁迫下梨枣光响应特征的变化,找到最适合植物生长的水分含量,保证枣树的各项生命活动正常进行。
目前关于枣树的研究较多,主要集中在对枣树的栽培技术品种筛选、病虫害、滴水灌溉方式以及枣树修剪等方面的研究,而对于水分胁迫对枣树光响应特征的影响研究较少[4]。陕西作为红枣的一大生产基地,为了充分了解枣树对干旱处理的响应,分别测定水分胁迫和对照处理下梨枣的净光合速率,并计算各项光响应参数,进行比较分析,探讨梨枣叶片对水分胁迫的适应变化,为梨枣的水分管理提供理论依据,推动陕北枣业的可持续发展。
1 材料和方法
1.1 材料
试验材料为3年生梨枣幼苗,由江苏新安果树公司提供。
1.2 研究方法
1.2.1 材料培养
首先,对3年生梨枣进行促根处理。将萘乙酸和吲哚丁酸(7∶3)混合加入8 L 1/2完全营养液中,配成促根溶剂,浸泡枣树20 h后进行室内盆栽,以沙粒为栽培基质,(室温25℃、光照环境),将土壤相对含水量控制在田间持水量的80%以上,3年生梨枣共栽植9桶,每桶各栽植3株幼苗,观察枣树的生长状况,待叶片萌发以后,对其进行控水处理。
1.2.2 试验设计
待梨枣叶片长大后,对其进行不同程度的水分处理,设置3个水分梯度,分别为正常水分处理(土壤含水量为田间持水量的80℅~85%,CK)、轻度水分胁迫(70%~75%,T1)、严重水分胁迫(40%~45%,T2),每一个土壤水分处理设置3个重复,每天同一时间使用电子台秤进行称重,以便及时补充当天所失水分。水分胁迫一周后,选取同一高度、相同部位、生长良好、长势一致的3年生梨枣叶片,运用便携式光合作用-荧光测量系统测定梨枣的净光合速率(Pn)。
1.3 测量方法
1.3.1 光响应曲线的测定
水分胁迫7 d以后,在自然CO2浓度条件下,使用便携式光合-荧光测量系统测定梨枣叶片的光响应参数,测量时选择3片健康、成熟的叶片作为试验材料,利用便携式光合-荧光测定系统的LED光源控制叶室内的光合有效辐射(PAR),分别设定为0、50、100、150、200、400、600、800、1000、1200、1500、1800 μmol·m-2·s-1,在不同光照强度下观测并记录净光合速率[5,6]。测量期间使叶片在每个光合有效辐射水平下适应120 s后观察并记录数据。
1.3.2 光响应曲线的测定
运用光合计算4.1.1软件计算光响应曲线的各项生理参数,其中非直角双曲线模型(Thornley,1976)的表达式为:
An(I)=(αI+Amax-Sqrt(αI+Amax)(αI+Amax)-4θαIAmax))/2θ-Rd
式中,An(I)为净光合速率,I为光强,θ为曲线的曲率,α为植物光合作用对光响应曲线在I=0时的斜率,即光响应曲线的初始斜率,也称为初始量子效率,Amax为最大净光合速率,Rd为暗呼吸速率[7,8]。
1.4 数据处理与统计分析
数据结果采用SPSS软件(版本21.0)和Microsoft Excel 2010进行光响应曲线的回归分析、单因素方差分析,使用光合计算4.1.1软件进行数据处理。各处理的比较采用最小显著差数法(LSD)进行多重比较。
2 结果与分析
2.1 不同水分胁迫下梨枣幼苗的光响应曲线
植物的光响应曲线衡量的是植物对光能的利用能力的指标[9]。如图1所示,在不同的水分条件下,梨枣叶片的Pn-PAR曲线的变化趋势基本保持一致。即不同水分处理后,梨枣叶片的Pn随着PAR的增加呈现出先增加后趋于平缓的趋势。当PAR<1000 mol·m-2·s-1时,梨枣叶片的Pn随着PAR的增加逐渐增加,涨幅很大;当PAR>1000 mol·m-2·s-1时,梨枣叶片的Pn变化趋势较慢,逐渐趋向于平缓。从曲线的变化幅度来看,水分胁迫条件下与CK相比有明显的差别,根据其胁迫水平的不同,呈现显著差异。具体来说,当PAR相同时,正常水分处理梨枣叶片的Pn增长速度最快、幅度最大。当PAR>200 mol·m-2·s-1时,轻度水分胁迫(T1)的梨枣叶片的Pn-PAR曲线接近CK曲线但总体低于CK,重度水分胁迫下Pn值较小,曲线较平缓。当PAR增大到一定的数值以后,各处理下Pn变化幅度很小。由此可以说明,水分胁迫处理对梨枣叶片的光响应过程有抑制作用,且随干旱程度的加深,Pn随PAR的变化幅度越来越小,梨枣对光能的利用能力越来越弱,不利于枣树叶片进行光合作用。
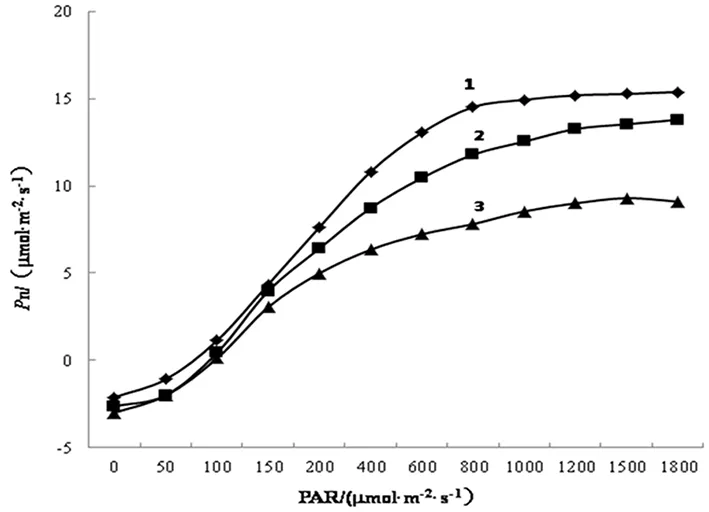
1.CK 2.T1 3.T2
2.2 不同水分胁迫对梨枣幼苗最大净光合速率、暗呼吸速率、表观量子效率、光饱和点和光补偿点的影响
在一定的环境条件下,最大净光合速率(Pnmax)反映的是植物叶片的最大光合能力[10]。由表1可知,在不同的水分处理条件下,梨枣幼苗的Pnmax随着水分胁迫的加剧逐渐降低,即最大光合能力逐渐下降。与对照组(CK)相比较而言,轻度水分胁迫下梨枣叶片的Pnmax下降程度不明显,二者之间无显著性差异,而重度水分胁迫下梨枣的Pnmax显著下降,与对照组相比降低了31.44%。表明重度水分胁迫下梨枣的光合作用受到显著影响,抑制了梨枣叶片最大光合能力的有效发挥[11],说明梨枣对水分亏损较敏感,无法适应干旱的环境条件。
暗呼吸速率(Rd)反映的是植物在黑暗条件下的呼吸速率,与叶片的生理活性有关[12]。由表1可知,梨枣的Rd随着土壤含水量的减少而不断增大,不同胁迫之下的暗呼吸速率差异显著。由此可知,不同程度的水分胁迫都会使叶片的暗呼吸速率升高,使得植物的呼吸作用增强,叶片生理活性增强,则植物对光合产物的消耗将会增加。有关研究表明:若植物的暗呼吸速率小,则说明植物在低Pn条件下更有利于维持碳平衡,即在低光照条件下,梨枣在适宜的土壤环境中更容易存活下来而不受影响。
表观量子效率(AQY)是光合作用中光能转化效率的指标之一,是净光合速率与相应光量子通量密度的比值[13],该值越高,光能转换效率就越高。由表1可知,根据其差异性水平发现,梨枣的AQY在正常水分处理和水分胁迫条件下均未发生显著性变化,轻度水分胁迫和重度水分胁迫下AQY分别下降了0.19%、4.31%,变化幅度很小,光能转化效率较低。根据表观量子效率逐渐变小的趋势可以得出水分胁迫几乎不影响梨枣叶片在弱光下吸收、转换和利用光能的能力,但根据有关研究,若植物出现水分亏损或水分过多的现象,都会致使植物的AQY降低[14],但针对不同的植物,仍然存在着显著差异,但在本研究中梨枣的AQY对水分亏损的反应较不明显。
光饱和点(LSP)指的是是植物利用强光的能力,LSP越高,表明植物利用强光能力越强且受到强光刺激时不易被抑制。光补偿点(LCP)反映的是植物利用弱光的能力,该值越小表明利用弱光的能力越强[15]。由表1可知,随着水分的减少,梨枣的LSP呈现逐渐减少的趋势,LCP则表现为逐渐升高的趋势,即土壤水分的减少使梨枣叶片利用强光和弱光的能力均减弱。梨枣在正常水分条件下LSP的数值较高,LCP的数值较低,表明三年生梨枣对光有较强的生态适应能力[16],所以梨枣幼苗在正常供水的条件下对光有较强的适应能力。综合分析LSP和LCP,表明随着土壤含水量的减少,梨枣进行光合作用的有效辐射范围越来越小,不利于梨枣进行光合作用。只有合适的土壤含水量,才能使梨枣对光的生态适应能力更好。

表1 不同水分处理下梨枣的光响应曲线特征参数值
注:表中小写字母表示P<0.05时各处理的差异显著性。
3 讨论
光合作用是植物体生长发育的重要生理过程,它的强弱对植物的生长发育及抗逆性有重要影响[17]。本文通过光响应曲线的拟合分析三年生梨枣的光合效率,并获得植物光响应特性的相关生理参数,但是研究植物光响应曲线的模型有许多,选用不同的模型提取的光响应参数和各项指标都存在差异[18]。本文仅仅选用非直线双曲线模型对水分胁迫下梨枣的光响应过程进行模拟,并以此为基础,通过计算获得光响应特征的各项相关参数。
本试验结果显示:不同水分胁迫下,梨枣的Pn-PAR曲线均呈现二次曲线的形式,且随着PAR的增大,Pn逐渐增大,当梨枣的PAR达到LSP后,光响应曲线的变化减慢,逐渐趋向于平缓。比较不同水分条件下Pn-PAR曲线的差异性可知,在光照强度较弱时,梨枣叶片的Pn差异不大;而PAR>200 μmol·m-2·s-1以后,Pn逐渐增大最后趋于平缓,Pn的大小顺序依次为正常水分处理>轻度水分胁迫>重度水分胁迫,表明在适宜的土壤水分环境中,梨枣叶片能较好的利用光能进行光合作用,随着PAR的增大,Pn逐渐增大,而水分胁迫下的梨枣叶片则较早就达到光饱和点,Pn不在继续增大。根据有关研究,植物的土壤含水率在60%以上时更有利于作物的生长,土壤含水率在田间持水率的60%以下时产生水分胁迫。由此说明重度水分胁迫对梨枣叶片光合作用的发挥有抑制作用,在同一PAR水平下,Pn随着胁迫的加深而逐渐降低,这与前人的研究保持一致[18]。
绿色植物的光响应特征的各项生理参数有利于我们研究植物的光合生理特性及植物生产力和对环境的适应性。在本试验中,随着水分胁迫的加剧,梨枣的Pn和LSP逐渐减小,Rd和LCP逐渐增加,而且在不同水分条件下存在显著差异(P<0.05),而AQY变化较小,有略微减小的趋势,但无显著性差异。由表1可知,在自然条件下(CK),三年生梨枣的Pnmax最大,具有较高的LSP和较低的LCP,是最适宜3年生梨枣生长发育的土壤水分条件。由此表明,在正常土壤含水量下最适合枣树的生长发育,水分胁迫促使梨枣叶片的光合能力下降,只有在正常灌溉的条件下,梨枣的光合能力最强,更有利于植物进行光合作用。根据王颖等人的研究来看,梨枣较适宜的灌溉量为田间持水量的45%,是陕北地区梨枣花期叶片进行光合作用的适宜水分含量,所以对于梨枣不同时期较适宜的灌溉量仍然有待于进一步的研究。
4 结论
综上所述,水分胁迫对3年生梨枣的光响应特征有影响,根据土壤含水量的不同,梨枣对其的反应也有很大差别。因此,在水资源充足的条件下,通过适当的增加土壤含水量,不仅不会影响梨枣的生理代谢过程,反而更有利于梨枣树的生长发育,提高枣树的产量。但本试验仍存在些许问题,本文采用非直角双曲线模型研究梨枣的光响应特征,由此得到的结论从科学的角度来看明显有许多不足,要想得到全面可靠的结论还需要进一步的研究,比如直角双曲线模型、非直角双曲线模型、指数模型很适合拟合正常生境下植物的光响应过程,使用这些模型对不同水分胁迫下3年生梨枣的光响应过程进行拟合,比较不同模型的拟合效果,结合实际情况,选择最佳模型,进行进一步的深入研究。
猜你喜欢
今日农业(2021年10期)2021-11-27
河北果树(2020年1期)2020-02-09
新课程·下旬(2017年7期)2017-08-14
Coco薇(2016年5期)2016-06-03
考试周刊(2016年6期)2016-03-11
少儿科学周刊·儿童版(2015年10期)2015-11-07
小说月刊(2015年10期)2015-04-23
西北林学院学报(2015年5期)2015-01-03
植物营养与肥料学报(2011年2期)2011-10-26
植物营养与肥料学报(2011年2期)2011-10-26
