
不同林龄麻栎林地下部分生物量与碳储量研究
2019-12-31 08:02:50胡海波张世豪卢洪霖
生态学报 2019年22期
王 霞,胡海波,*,张世豪,卢洪霖
1 南京林业大学南方现代林业协同创新中心,南京 210037 2 江苏省水文水资源勘测局镇江分局,镇江 212100 3 江苏省水土保持与生态修复实验室,南京 210037 4 江苏省句容市林场,句容 212424
森林碳汇具有成本低、易执行、潜力大、见效快等特点,已被世界各国学者所认同,并进行了积极探索,取得了丰富成果。森林生态系统是全球重要碳库,储存了全球陆地生态系统2/3以上的有机碳,分别储存了陆地生态系统地上部分与地下部分有机碳的80%和40%[1-2]。林分组成通过调节有机碳在地表植被、乔木层等不同有机碳库间的分配比例,影响整个森林生态系统碳储存功能[3-4],森林植被碳储量直接反应了森林生态系统的碳汇潜力,是研究森林生态系统与大气圈之间碳交换的基本参数。
森林生物量在评价林分生产力、结构与功能,研究森林物质循环与能量流动等方面发挥着重要作用[5-6],同时是森林生态系统碳储量的重要数据来源和全球碳循环的重要构成因子。生物量的研究方法有样地实测建模法、生理生态模型法、遥感信息模型法等[7]。相对生长方程模型法是样地实测建模法常用的生物量模型,其常用的变量通常为易测的林分调查因子,如胸径(D)和树高(H),此外还有学者添加树龄、树冠、海拔等因子,这不仅增加了野外调查的工作量和难度,还加大了模型估算的复杂程度,进而影响模型的应用。大部分研究者以胸径(D)作为唯一自变量或胸径(D)与树高(H)两个测树因子作为自变量而建立单木生物量模型,再根据每木检尺的资料估算林地乔木生物量。相对生长方程模型通常包含直线回归、对数回归、逆矩阵式、二次方程式、三次方程式、复合曲线式、幂回归式、“S”形曲线式、生长曲线式、指数回归式、Logistic曲线式11种数学回归模型。通过构建森林生物量模型不仅可以减少野外调查与测算的工作量,减轻对植被、土壤的破坏,防止水土流失,还能准确估算森林生物量,且可在相似森林类型中利用已有的模型预估其生物量,因此森林生物量模型的构建是极其必要的。
目前森林生物量的研究多集中在地上部分,因地下部分根系难以获取收集,故有关地下部分的研究相对较少,而树木根系在森林生态系统物质和能量循环过程中起着中介作用,且在输送营养物质、保持肥力和水土及固碳等方面具有重要的意义,生态系统中根系生物量占比较大,其生物量占总生物量的10%—20%[8-9]。由于根系深埋地下,研究方法与过程繁琐,且不易观察和测量,往往存在较大误差,因此最初的研究相对薄弱,二十世纪中期以来,森林根系逐渐成为研究热点,且研究日趋成熟,森林生态系统的功能趋于完善和深入[10]。
麻栎(Quercusacutissima),又名橡树,是壳斗科(Fagaceae)、栎属(Quercus)落叶乔木树种,广泛分布于暖温带和亚热带地区[11]。麻栎常见于次生林中,具有耐干旱、耐贫瘠、根系庞大的特点,是荒山瘠地的先锋树种和水土保持的优良树种;又栎炭热值高、无烟、碳素含量高,是理想的生活、工副业燃料和工业原料[12]。目前,国内对麻栎的研究主要集中在栽培技术[13-15]、经营措施[16-17]、薪炭林与人工林碳循环[18-20]等方面,且集中于地上部分,对根系研究较少,因此本文以不同林龄麻栎林为研究对象,采用相对生长方程模型法,构建麻栎根系生物量回归模型,估算麻栎林根系碳储量,以期为麻栎林的经营管理及碳汇功能评估等提供技术支撑。
1 研究区概况
试验地位于江苏省句容市下蜀林场和句容林场(119°12′E,32°07′N),处于宁镇山脉低山丘陵地带中段的空青山,属北亚热带季风气候区,四季分明,光照充足。地形是丘陵山地,起伏较平缓。年平均气温15.2℃,年均降水量1055.6mm,年际变化较大;年均相对湿度79%;无霜期233d。土壤以黄棕壤为主。该区森林植被属我国北亚热带东部区,是北亚热带向暖温带过渡的植被类型,地带性植被为带有常绿成分的落叶阔叶混交林。试验地是以麻栎为主的天然次生林,灌木植物主要有牛鼻栓(Fortuneariasinensis)、华紫珠(Callicarpacathayana)、枸骨(Ilexcornuta)、白檀(Symplocospaniculata)、野蔷薇(Rosamultiflora)等,草藤本植物主要有爬山虎(Parthenocissustricuspidata)、铁线蕨(Adiantumcapillus-veneris)、石血(Trachelospermumjasminoides)等。
2 研究方法
因朱燕飞等[21]已研究过相同林分地上部分的生物量,本文仅探讨麻栎林地下部分根系的生物量与碳储量。
2.1 样地设置与采样
在江苏省句容市的下蜀林场和句容林场,选取幼龄林(12a)、中龄林(35a)、近熟林(47a)和成熟林(60a)的麻栎次生林,每个林分按坡位各选择3个调查样地(20m×20m),共12块样地进行调查,对样地进行每木检尺,测量胸径、树高与郁闭度等指标(表1),确定标准木,并调查样地的灌木郁闭度、草本植物覆盖度(表1)以及土壤性质等环境因子(表2)。
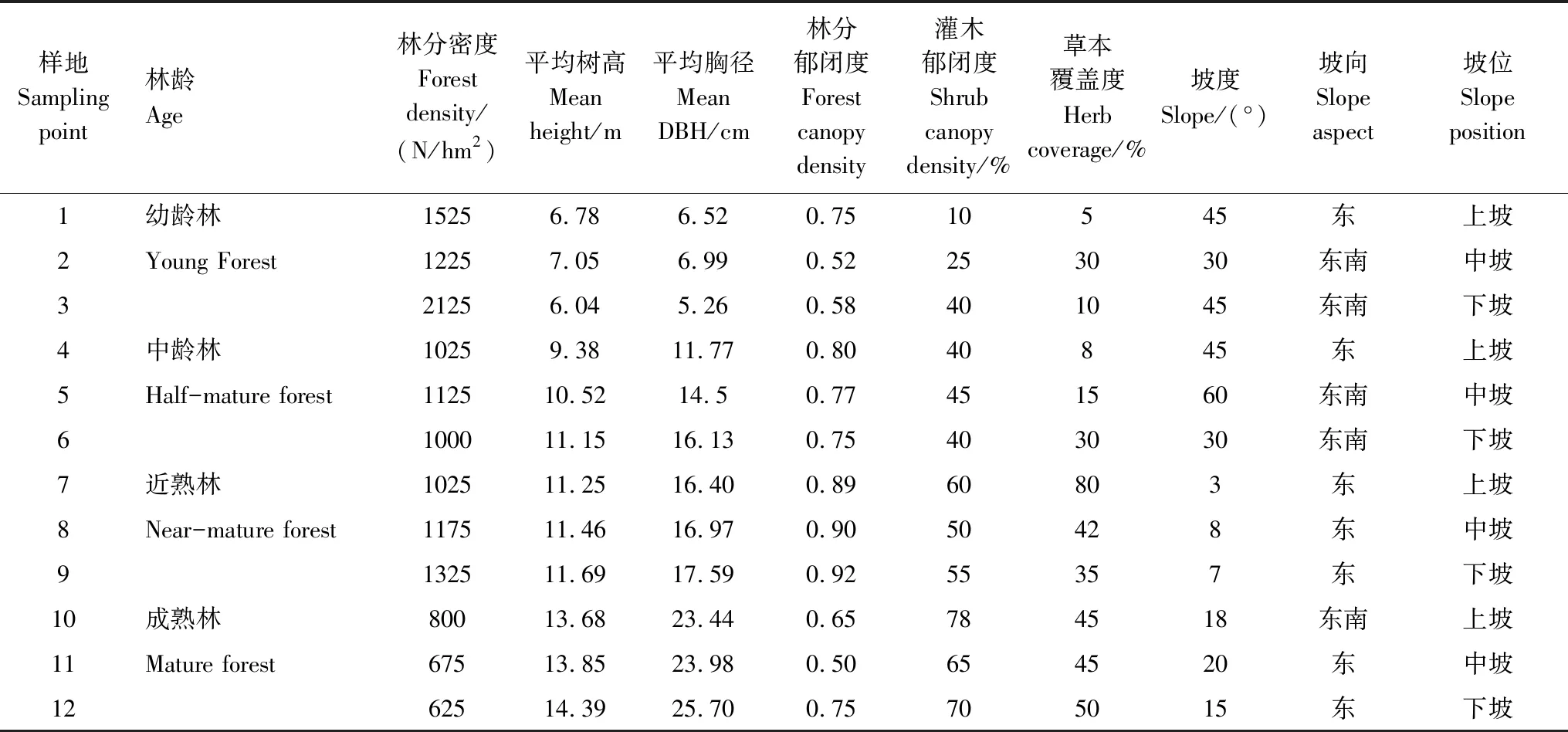
表1 试验地基本情况 Table 1 General conditions of experimental forest stands
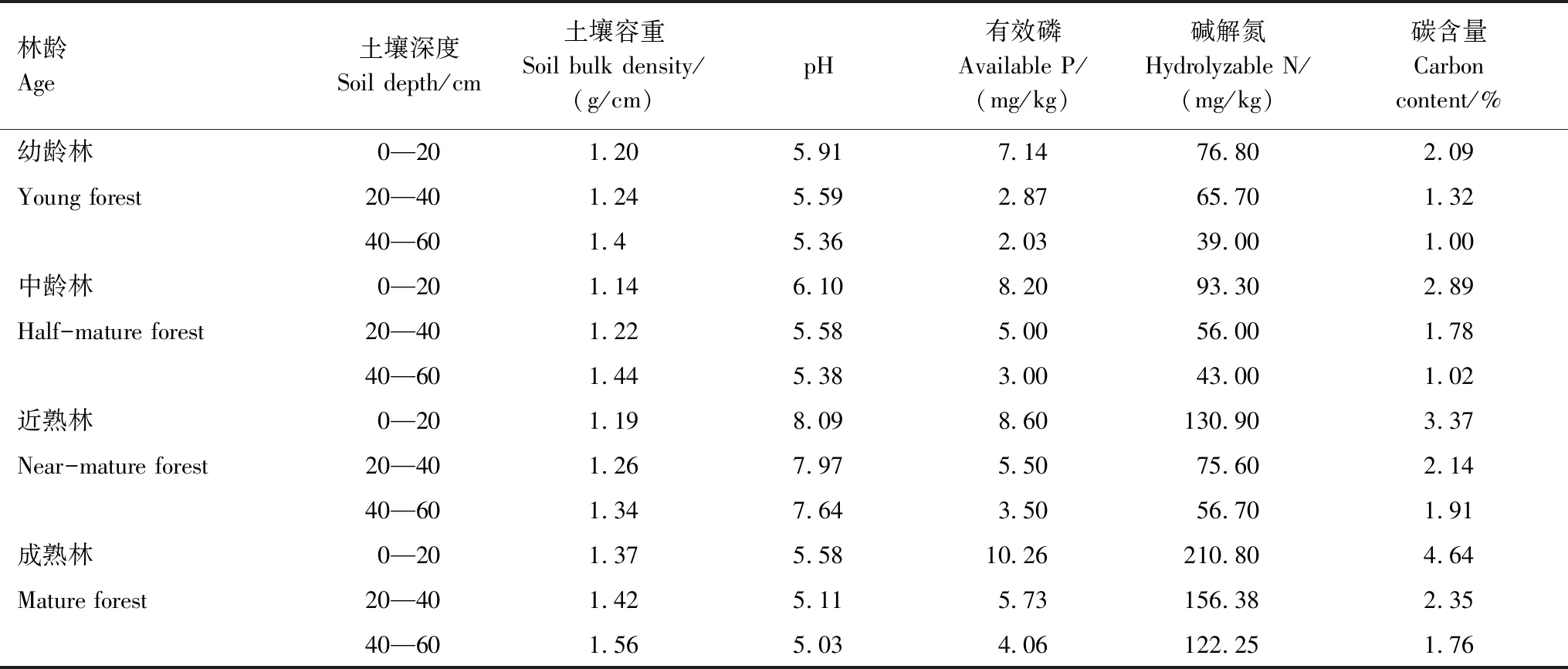
表2 麻栎林土壤基本理化性质Tab 2 Basic physical and chemical properties of the soil in Quercus acutissima forest
乔木层样品采样:为了减少对森林的破坏,在每个样地内各选择2棵标准木,共计24棵标准木,采用全根挖掘法,将每个样地标准木伐倒,挖出全部根系,按照根桩(实体部分)、粗根(>2cm)、大根(0.2—2cm)、细根(<0.2cm)分级收集,分别测定鲜重,各级根系样品均采集500—1000g,带回实验室处理。
灌草层样品采样:在每个样地内沿对角线分别在坡上、坡中、坡下,各设置三个2m×2m的灌木样方、1m×1m的草本样方,记录每个样方内的植物种类,收集样方内的全部灌、草植物的根系并测定鲜重,取500—1000g样品,带回实验室进行处理。
2.2 样品处理及测定方法
收集到的样品去除杂物后称重记录,再用超纯水洗净,于70—80℃烘箱中烘干至恒重后称重记录,测定含水率,以计算生物量。将烘干的样品剪碎,用高速粉碎机磨细后,做好标记袋装保存,用于实验分析。根系碳含量的测定采用重铬酸钾氧化-外加热法[22]。
2.3 数据处理与分析
根据式(1)计算麻栎林乔木层、灌草层根系碳储量,并根据野外调查结果和室内分析数据,用Excel进行数据处理,用SPSS 22.0对数据进行相关性分析,并建立回归模型。
碳储量(t/hm2)=生物量(t/hm2)×碳含量(%)×10-2
(1)
3 结果与分析
3.1 麻栎根系生物量
3.1.1单株麻栎根系生物量回归模型建立
以胸径(D)、胸径-树高(D2H)分别为自变量,用直线回归、对数回归、逆矩阵式、二次方程式、三次方程式、复合曲线式、幂回归式、“S”形曲线式、生长曲线式、指数回归式、Logistic曲线式11种数学回归模型,拟合出11种模型的各级根系生物量与其总生物量的回归方程,各回归方程经检验均达到显著水平(附件1)。通过比较发现,幂回归方程的拟合效果最好(表3),该模型除细根外均具有很好的拟合结果,与朱燕飞等对相同麻栎林地上部分的生物量模型具有相同的结论[21]。
由表3可知,在方程W=aDb中,根桩、粗根、大根回归方程的相关系数都在0.98以上,粗根的生物量相关系数更是达到了0.996,是各级根系中相关系数最高的。方程W=a(D2H)b拟合结果与方程W=aDb的相似,相关系数差值在0.001—0.003之间。两个幂回归方程细根生物量相关系数均小于0.8,这可能是由于细根受土壤理化性质影响较大,以及根系较小难以发现,故导致拟合结果产生偏差。

表3 单株麻栎各级根系生物量回归方程 Table 3 Regression equations for individual root biomass at different root levels
由表3知,方程W=a(D2H)b与W=aDb的拟合结果相似,因此这两个方程均适用于麻栎单株林木根系生物量估算,本文选择方程W=aDb,以与朱燕飞对同一麻栎林地上部分生物量模型的研究一致。由此,建立单株麻栎根系生物量W与胸径(D)的回归方程,即:
W=0.434D1.507+0.247D1.515+0.352D0.735+0.726D0.187
(2)
3.1.2麻栎林根系生物量及其分配
根据单株生物量方程(式2)及标准地资料,参考马钦彦[23]的林木生物量估算法(式3),可得到麻栎林根系生物量模型(式4),公式如下:
(3)
(4)
式(3)、式(4)中,W为单位面积生物量(t/hm2),k为径阶组数,Di、ni分别表示第i径阶组的平均胸径(cm)和株数,a、b为参数。
运用麻栎林根系生物量模型(式4)及林分径阶资料,计算得到不同林龄麻栎根系生物量(表4)。由表4知,幼龄林、中龄林、近熟林、成熟林麻栎根系生物量分别为14.81t/hm2、41.15t/hm2、50.36t/hm2、53.75t/hm2。麻栎各级根系生物量呈现出一定规律性,其大小顺序为根桩>粗根>大根>细根,根桩生物量占据了主导地位,体现出其有机质的积累水平,同时发现随根系的生长发育,细根生物量逐渐减小。根系生物量随林龄增大而增大,但大根与细根生物量在达到近熟林期后开始下降,且近熟林和成熟林根系生物量相近,这可能是因为幼龄林到近熟林成长过程中需要大量细根吸收营养物质以维持其正常生长。成熟林根系生物量分别是幼龄林、中龄林、近熟林的3.63、1.31、1.07倍,说明幼龄林成长到近熟林根系生物量积累迅速,而近熟林到成熟林生长过程中生物量积累缓慢。
从各级根系生物量分配比来看,随林龄增大,根桩、粗根生物量占总根系的比例也在增大,根桩生物量占比均在50%以上,粗根生物量占比在30%以上;而大根与细根则相反,这是因为麻栎在幼龄林期到近熟林期过程中生长迅速,需大量大根、细根吸收营养物质维持其生长,而在近熟期到成熟期过程中,生长缓慢趋于稳定,且细根的周转率通常在0.5—1.2次/a[24],因此大根与细根生物量占比随林龄增大而减小。随着林龄增大,根桩生物量占比从51.87%增加到59.43%,粗根生物量占比从29.98%增加到34.68%,大根生物量占比从10.11%减小到4.29%,细根生物量占比从8.04%减小到1.60%。
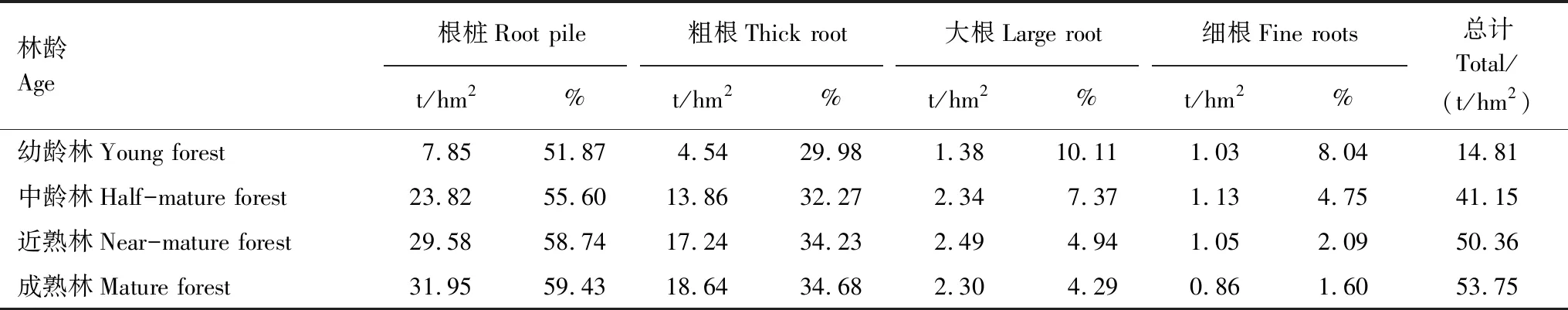
表4 不同林龄麻栎林各级根系生物量 Table 4 Biomass of different root levels of Quercus acutissima for different stand ages
3.1.3麻栎群落根系生物量结构
从表5可以看出,灌木根系生物量从幼龄林的0.48t/hm2增加到成熟林的1.71t/hm2;草本植物根系生物量从0.13t/hm2增加到成熟林的0.60t/hm2。麻栎群落根系生物量随林龄的增大而增大,不同林龄麻栎群落根系生物量为15.42—56.06t/hm2,成熟林根系生物量分别是幼龄林、中龄林、近熟林的3.64倍、1.34倍、1.08倍。麻栎幼龄林群落根系生长快速,到近熟林期群落根系生物量大量积累,近熟林与成熟林群落根系生物量接近,说明麻栎林达到近熟林期后群落根系生物量积累缓慢。
同时看出,各林层根系生物量大小顺序为:乔木层>灌木层>草本层,且均随林龄的增大而增大。不同林龄麻栎群落根系生物量均以乔木层最大,占林分地下根系生物量的比例达到95%以上;占比最小的是草本植物,均在1.1%以下;灌木根系生物量占比为1.37—3.14%。幼龄林处于生长初期,林分郁闭度小,为灌木生长提供了良好的条件,因此幼龄林灌木根系生物量均比其他林龄所占的比例略大;草本植物根系生物量无明显变化规律。

表5 不同林龄麻栎林群落根系生物量 Table 5 Root biomass of Quercus acutissima community
3.2 麻栎群落根系碳含量
由表6可知,从乔木层来看,麻栎各级根系碳含量大小顺序为:根桩>粗根>大根>细根。根桩最大碳含量为近熟林45.12%,最小为幼龄林41.26%;粗根最大碳含量为中龄林的42.44%,最小为成熟林的40.26%;大根最大碳含量为中龄林的42.36%,最小为成熟林的40.01%;细根最大碳含量为近熟林的40.02%,最小为成熟林的38.81%,可以看出麻栎各级根系碳含量存在一定差异,但都低于碳汇研究中常采用的50%;随林龄增大,各级根系碳含量并未随之增大,而是呈先增大后减小的规律,且不同林龄之间麻栎根系碳含量差异不显著,平均碳含量为37.42%—42.43%。麻栎林灌木根系碳含量为41.84%—43.79%,草本植物根系碳含量为34.03%—38.48%,不同林龄麻栎林灌木层和草本层根系碳含量无明显变化规律。
3.3 麻栎群落根系碳储量
3.3.1麻栎林根系碳储量
由表7可知,麻栎林根系碳储量随林龄增大而增大。幼龄林、中龄林、近熟林、成熟林根系碳储量分别为6.01t/hm2、17.41t/hm2、21.79t/hm2、21.99t/hm2,且成熟林根系碳储量分别是幼龄林、中龄林、近熟林的3.66倍、1.26倍、1.01倍,近熟林与成熟林的碳储量接近,表明麻栎根系到近熟林期后碳素积累缓慢,且近熟林郁闭度大,生长空间小,麻栎根系生长趋近缓慢,故而碳素积累缓慢。可知,随林龄的增大,麻栎林根系生物量不断增加,其碳储量也随之增加。
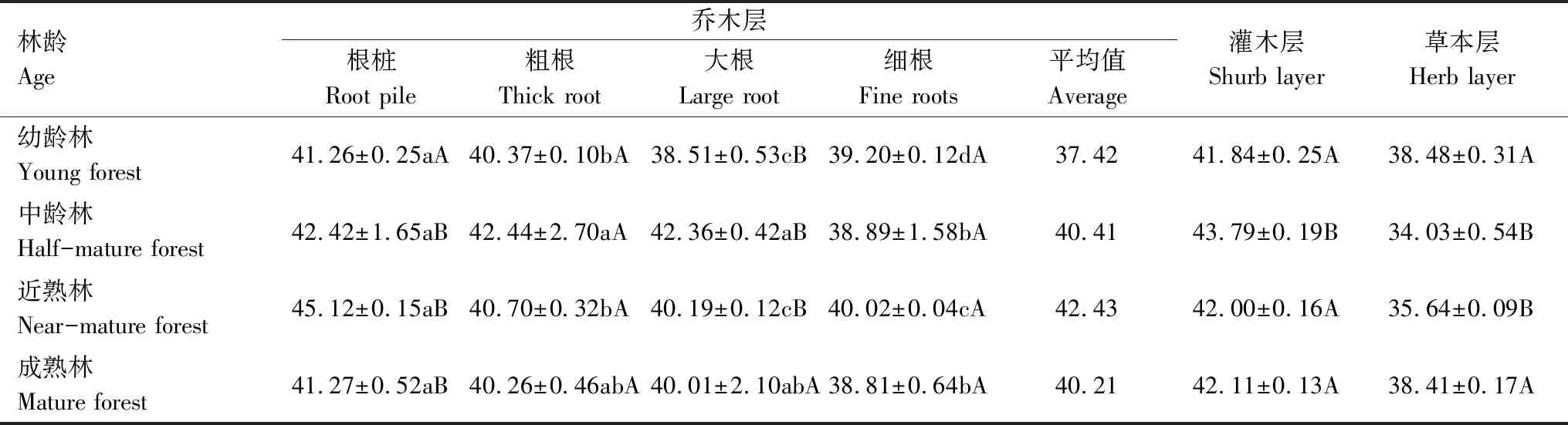
表6 不同林龄麻栎林各级根系碳含量/%Table 6 Carbon content of different root levels of Quercus acutissima at different stand ages
表中数据为均值±标准误差,数据后不同小写字母(a、b、c)表示同一林龄不同等级根系、不同大写字母(A、B)表示同一等级根系不同林龄在0.05水平上的差异显著性,字母相同表示差异不显著(P>0.05),字母不同表示差异显著(P<0.05)
各级根系碳储量大小顺序为根桩>粗根>大根>细根。细根碳储量所占比例最小,细根占根系碳储量的最大值为6.72%;根桩所占的比例最大,最大值为61.27%,表明根桩是麻栎林地下根系碳储量的主体。根桩与粗根碳储量占比随林龄增大基本呈增大的趋势,而大根与细根碳储量占比随林龄增大而减小,与根系生物量的变化规律一致。

表7 不同林龄麻栎林各级根系碳储量Table 7 Carbon storage of different root levels of Quercus acutissima at different stand ages
3.3.2麻栎群落根系碳储量
由表8知,麻栎群落根系碳储量随林龄的增大而增大,且各林龄碳储量均表现为乔木>灌木>草本,幼龄林、中龄林、近熟林、成熟林群落根系碳储量分别为6.26t/hm2、17.74t/hm2、22.37t/hm2、22.94t/hm2,成熟林群落根系碳储量分别是幼龄林、中龄林、近熟林的3.82倍、1.29倍、1.03倍,近熟林与成熟林群落根系碳储量接近,麻栎林群落近熟林期后碳储量增加缓慢,与生物量变化规律一致。
从各林分层次来看,乔木根系碳储量随林龄增大而增大,其占林分地下部分根系总碳储量的比例均在95%以上,与生物量变化规律相似,因此乔木层根系是麻栎群落地下部分根系的主要碳汇场所。灌木根系碳储量也随林龄增大而增大,幼龄林、中龄林、近熟林、成熟林灌木根系碳储量分别为0.20t/hm2、0.25t/hm2、0.36t/hm2、0.72t/hm2,其中幼龄林碳储量所占比例最大,为3.24%,与生物量变化规律一致。草本植物根系碳储量同样随林龄增大而增大,幼龄林、中龄林、近熟林、成熟林草本层根系碳储量分别为0.05t/hm2、0.07t/hm2、0.22t/hm2、0.23t/hm2;草本植物根系碳储量所占比例最小,在1.01%以下。

表8 不同林龄麻栎群落根系碳储量Table 8 Root carbon storage of Quercus acutissima community at different stand ages
4 讨论
森林生物量通常采用解析木法直接测定与生物量模型法间接测定,解析木法对森林破坏严重且较复杂,而生物量模型法简便快捷且对森林环境影响小。通常以胸径(D)、胸径-树高(D2H)为自变量建构模型,宋博等[25]的研究表明仅含有胸径的生长方程是一种最为简单的模型形式,且具有较高的预测精度。本文以易测得的胸径与树高测树因子为自变量,采用相对生长方程模型法,通过11种回归方程比较,得出W=aDb与W=a(D2H)b两个幂回归方程拟合结果最好,且相关系数差异不大,为与前人对同一麻栎林地上部分生物量模型的研究[21]一致,本文选择W=aDb作为麻栎根系生物量模型,弥补了以往只对全根生物量估算而误差大的问题,也为今后麻栎林生物量研究提供了依据。
麻栎各级根系生物量大小顺序是:根桩>粗根>大根>细根,根桩生物量占据主导地位,与许雯对马尾松林根系生物量研究结果一致[26]。麻栎幼龄林、中龄林、近熟林、成熟林群落根系生物量随林龄的增大而增大,影响生物量的因素有郁闭度、林分密度、立地条件等,由于麻栎幼龄林期郁闭度小、生长空间大、光照充足,有利于光合作用积累有机物,因此幼龄林到近熟林根系生物量增长迅速,而近熟林期郁闭度大、光照不足,根系生物量增速缓慢且与成熟林接近。
麻栎各级根系碳含量大小顺序为:根桩>粗根>大根>细根,差异显著,但都低于国际上常用的0.5的含碳系数[27],由于国际含碳系数是不分树种、不分林龄的[28],且植物既有高碳组织又有低碳组织,我国阔叶树平均含碳系数大多不足0.5,而针叶树平均含碳系数大多高于0.5[29],同时存在立地条件、气候条件以及物种差异性,所以本研究麻栎各级根系碳含量低于50%。因此,若采用国际上含碳系数估算麻栎林碳储量,将产生较大的系统误差,这也反映出采用国际含碳系数有一定的局限性与不确定性。麻栎群落灌木和草本植物根系碳含量并无明显变化规律。
麻栎幼龄林、中龄林、近熟林、成熟林群落根系碳储量随林龄的增大而增大,幼龄林、中龄林碳储量增速快,而近熟林较慢,由于我国人工幼、中龄林多,可知中国森林的碳汇功能将进一步增强[30]。各林龄群落碳储量均表现为乔木层>灌木层>草本层,且乔木层根系碳储量占群落地下部分根系总碳储量的比例在95%以上,表明乔木层根系是群落地下部分根系碳汇的主要场所,且随林龄增大而增大,说明随林龄增长麻栎根系碳素在逐渐积累,与不同林龄麻栎林地上部分碳储量变化规律一致[21]。
麻栎群落地下部分根系生物量与碳储量均随林龄的增大而增大,幼龄林到近熟林生长过程中根系生物量与碳储量增加快速,近熟林后根系生物量、碳素积累缓慢且与成熟林接近。因此在森林经营管理以及碳汇管理时,从麻栎林根系生物量与碳储量的角度出发,同时考虑生态、经济效益,在不同林龄期控制和减少森林采伐,延长轮伐期,成向荣[31]等的研究表明间伐可以提高林分生物量与碳储量,因此还可通过增大间伐强度来促进森林树木生长,实现林分结构的调整,从而增强森林的碳汇功能,促进森林的可持续发展。
猜你喜欢
广东蚕业(2022年4期)2022-11-23 13:31:55
江西农业大学学报(2022年1期)2022-02-04 11:51:42
江西农业大学学报(2021年2期)2021-05-16 02:18:24
林业勘查设计(2021年2期)2021-04-08 08:27:42
土壤学报(2020年6期)2021-01-05 13:26:54
湖北农机化(2020年16期)2020-10-21 01:29:08
江西农业(2020年13期)2020-09-08 07:44:54
农民致富之友(2020年8期)2020-05-11 05:54:38
广东农业科学(2019年2期)2019-04-09 08:32:54
防护林科技(2016年5期)2016-09-05 01:19:50
