
陆地棉种质资源的表型及SSR遗传多样性分析
2019-12-25 08:43贾子昉王清连董娜
生物技术通讯 2019年5期
贾子昉,王清连,董娜
河南科技学院生命科技学院/现代生物育种河南省协同创新中心/河南省棉麦分子生态与种质创新重点实验室,河南新乡453003
开展棉花品种的遗传多样性研究具有重要的意义,近年来对自育品种、亚洲棉、陆地棉等的遗传多样性研究越来越多,对种质资源遗传多样性的研究是目前育种工作者的主要任务。目前,分子标记如随机扩增多态性DNA(random amplified polymorphic DNA,RAPD)、扩增片段长度多态性(amplified fragment length polymorphism,AFLP)、限制性内切酶片段长度多态性(restriction fragment length polymorphism,RFLP)、简单重复序列(simple sequence repeat,SSR)等已被广泛应用于棉花遗传多样性研究。其中,SSR 以其具有较高的多态性、共显性、DNA 样本用量少而被大范围应用[1-4]。前人利用表型性状对棉花遗传多样性进行了比较系统的研究[5-8],但是将表型性状与分子标记两者同时应用于遗传多样性研究的报道较少。本研究中,我们主要利用SSR 标记及表型分析探讨300 份种质资源的遗传多样性,为育种工作者提供丰富的品种遗传多样性信息,为合理进行亲本杂交选配,培育高产、优质和多抗新品种提供理论依据。
1 材料和方法
1.1 材料
供试的300 份陆地棉种质资源材料由河南科技学院生命科技学院的棉花种植试验田提供(表1)。试验材料于2016 年4 月种植于试验田,同时开始2 年的田间表型性状调查,室内试验于2018年10 月进行。
1.2 棉花基因组DNA 的提取
在棉花生长旺盛时期,每一个品种随机任取2 株上未展平的新鲜嫩叶1~2 片,放入预冷的离心管中,立即加入600 μL 预冷的DNA 提取液,用电钻法将嫩叶组织磨至细碎。用改良CTAB 法提取DNA[9-10]。DNA 溶解在灭菌的TE 中,浓度为50 ng/μL。
1.3 SSR 引物的筛选
随机选取16 份种质材料对372 对SSR 引物进行初筛,共得到72 对差异性引物。372 对SSR 引物序列信息来自棉花分子标记数据库(CMD;http://www.cottonmarker.org),由南京金斯瑞生物科技有限公司合成,包括84 对BNL 引物、20 对JESPR 引物、267 对NAU 引物、1 对TMB 引物。从72 对差异性引物中选取重复性好、条带清晰稳定、信号强的38 对引物(表2),在300 份供试材料间进行PCR 反应[11]。
1.4 PCR 扩增及电泳检测
PCR 反应体系为10 μL,包括1.0 μL 模板,正、反引物(5 μmol/L)各0.5 μL,5 μLTaqMasteMix(含有TaqDNA 聚合酶、PCR 缓冲液、Mg2+、dNTPs),3 μL ddH2O。扩增程序:95℃预变性2 min;94℃变性45 s,57℃退火45 s,72℃延伸60 s,循环30 次;72℃延伸7 min;10℃保存。扩增产物在北京六一仪器厂生产的DYY-6C 型稳压稳流电泳仪上经9%聚丙烯酰胺凝胶检测,恒定电压180 V,电泳1 h,银染后观察并照相[12-13]。
1.5 数据处理与分析
对2 年田间数据进行平均值处理,采用SPSS 19.0 统计分析软件对表型数据进行统计描述分析,得到方差、标准差、变异系数等参数,并进行
聚类分析。对SSR 电泳图谱上的条带,有带记为“1”,无带记为“0”,缺失记为“2”[9,14]。用NTSYS-pc2.10e 软件处理SSR 分型数据,计算品种对之间的遗传相似系数,并获得相似系数矩阵。用SHAN 程序中的UPGMA(Unweighted pair-group method with rithmetic averages)方法进行聚类分析,并通过Treeplot 模块生成聚类图。

表1 300份供试棉花材料
2 结果
2.1 300 份种质的表型性状遗传多样性方差分析
根据性状调查结果,对300 份陆地棉种质资源的生育期、单铃重、株高、衣分、铃数、皮棉产量、子棉产量、发病指数等产量性状进行统计分析(表3)。上述8 个主要农艺性状中,300 份种质资源的生育期、单铃重、株高、衣分、铃数、皮棉产量、子棉产量等性状都有极显著差异,只有发病指数不参与显著性分析;铃数、皮棉产量、子棉产量、发病指数等的变异系数很大,均超过20%;单铃重的变异系数大于10%;生育期、衣分、株高的变异系数均低于10%;8 个表型性状变异系数大小依次为发病指数>皮棉产量>铃数>单铃重>株高>衣分>子棉产量>生育期;300 份种质资源间的大部分表型性状存在显著差异,说明所选用的陆地棉种质资源的表型性状差异明显、变异范围大,具有比较好的代表性。对300 份棉花供试材料依据Shannon-Weaver 信息指数公式计算形态多样性指数,从结果可以看出300 份供试材料的平均多样性指数为5.597,8 个形态指标的多样性指数依次为5.703、5.696、5.700、5.699、5.633、5.608、5.622、5.117,均表现出较高的多样性;多样性水平依次为生育期>衣分>株高>单铃重>铃数>子棉产量>皮棉产量>发病指数。变异系数和Shannon-Wiener 多样性指数在表型性状方面,2 个计算方法所得结果并不一致,这与前人的研究结果相同。原因可能是因为多样性指数和变异系数的性质以及计算方法不同。Shannon-Wiener 多样性指数强调的是不同层次分布的均匀度,而变异系数强调的是数值变异的大小。
2.2 300 份种质的表型性状聚类分析

表2 38 对多态性引物信息
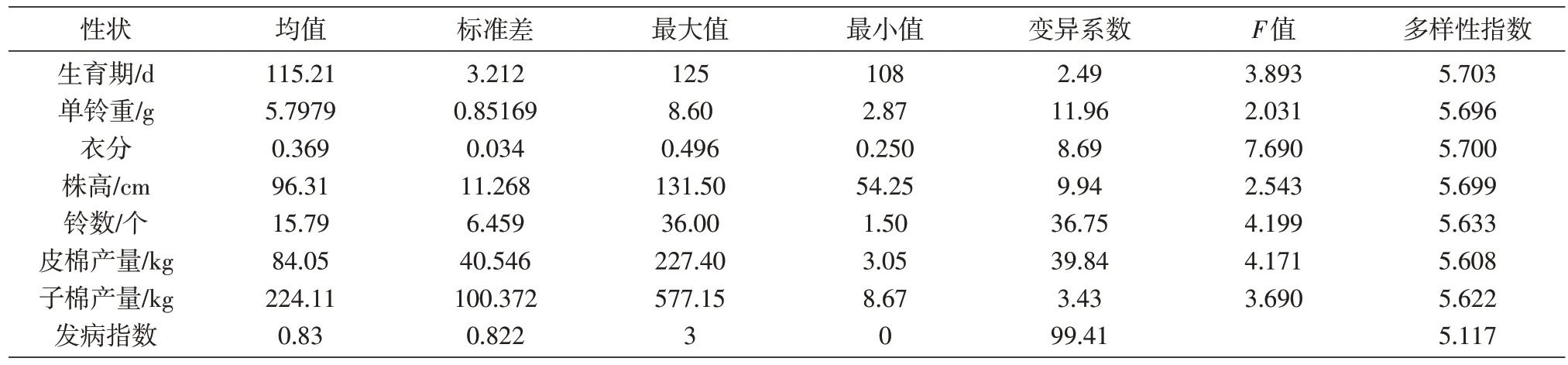
表3 300份种质资源表型性状差异
利用SPSS19.0 软件,根据调查的形态数据进行聚类分析,得到亲缘关系聚类图(图1)。可以看出,300 份种质资源被分为Ⅰ和Ⅱ两大类,Ⅰ包括151 份品种,Ⅱ包括149 份品种。Ⅰ和Ⅱ又各自被分成Ⅰ-1、Ⅰ-2 和Ⅱ-1、Ⅱ-2,它们又分成Ⅰ-1-A、Ⅰ-1-B、Ⅰ-2-A、Ⅰ-2-B、Ⅱ-2-A、Ⅱ-2-B 等类别。
Ⅰ-1-A1 主要是一些抗虫、抗黄萎病、中大铃的优质品种,包括75、104、188、272、57、198、72、133、288、135、267 等46 份材料。
Ⅰ-1-A2 包括79、169、12、94、146、176、4、42、50、69、168、286 等27 份材料,该类群的特征不太统一,主要是色泽比较差的品种,还有一些高蛋白耐盐优质品种、高抗黄萎病品种,如DES926;还有一个是国内引进品种,红桃。
Ⅰ-1-B1 包括148、268、144、278、172、237、242、256、277、128、56、250、282 等13 份材料,主要是一些创新种质及OLD 品种,生育期比较长,铃重居中,株高比较低,棉铃虫抗性较低。
Ⅰ-1-B2 包括183、293、190、255、35、187、129、232、260、67 等28 份材料。只有长德184、抗黄萎164 两个品种抗虫,其他均不抗虫;生育期都很长,株高低。
Ⅰ-2-A 包括88、197、95、155、159、58、140、249、264、248、269 等17 份材料。株高偏低,均为不抗虫品种。
Ⅰ-2-B1 包括283、284、36、231、234、139、161、38、292、285、296、280 等12 份材料,主要是一些丰产品种,不抗虫,大部分为国外引进品种。
Ⅰ-2-B2 包括48、127、138、257、167、126、273、136 等8 份材料。产量很低,铃数少,衣分低,不抗虫。
Ⅱ-2-A 包括142、264、201、62、130、65、212、60、173、37、195、73 等20 份材料,个别抗虫,整体生育期较长,矮杆,丰产。
Ⅱ-2-B1 包括218、241、131、238、253、30、177、243、270、281、107 等22 份材料。该亚类的特征是不抗虫。
Ⅱ-2-B21 包括24、109、157、196、90、215、74、125、118、91、96、5、6 等34 份材料。该类群的品种大部分高产,株高偏低。
Ⅱ-2-B22 包括251、294、14、244、290、3、236、97、210、233、77、240 等32 份材料。衣分高,抗黄萎病指数比较高。
2.3 SSR 标记多态性分析
在筛选出的38 对引物中,共检测出126 个等位位点,多态性位点为90 个,占71.43%,多态性比例较低。每个标记等位位点的数量范围为2~10 个,平均3.66 个;每个标记的多态性位点为1~6个,平均2.42 个;引物JESPR291 扩增出的多态性条带最多,为6 条,其次为引物NAU2679、NAU3888,均为5 条。
2.4 成对相似系数分析
利用NTSYS-pc2.10e 和Excel 软件计算品种对间的遗传相似系数,相似系数矩阵由于过大没有列出,仅列出品种间的相似系数统计分析表(表4)。结果显示,各品种对的相似系数为0.175~0.905,平均值0.533。遗传差异较大(相似系数小于0.500)的品种对占46.31%,比例适中。46%品种对的遗传相似系数较小,说明品种对之间的遗传多样性比较丰富。其中,M183 和M82 的遗传相似系数最小,为0.175,遗传差异最大;M101 和M81 的遗传相似系数最大,为0.905,遗传相似度极高。
2.5 SSR 标记的聚类分析
通过遗传相似系数进行UPGMA 聚类(图2),阈值为0.400 时,300 个品种被聚为Ⅰ和Ⅱ两大类群。类群Ⅰ包括150 份种质,阈值为0.590 时又被分为Ⅰ-1、Ⅰ-2 两大类。Ⅰ-1 在阈值为0.616 时被分为两类,Ⅰ-A 包括148 份种质,遗传相似系数为0.632~0900;Ⅰ-B 仅有1 份种质苏联棉91系。Ⅰ-2 仅包括1 个品种博乐07-301。
Ⅰ-A 在阈值为0.65 时被分为9 个亚类。Ⅰ-A1 包括1、22、21、9、71 等68 份种质,来源比较复杂,大部分为抗虫品种,产量高。Ⅰ-A2 包括6、15、116、41、123 等24 份种质,品种复杂,无明显的共同特征,但大体上大部分均为抗虫品种。Ⅰ-A3 只有库车T94-6、鲁农9648 两个品种。Ⅰ-A4 包括新陆早6 号、陕早2786、新陆棉201、徐州219、冀远12-13,为OLD,不抗虫品种。Ⅰ-A5 包括山农丰抗6 号、R01/中三都4121 F9、宿9108/R03 F9。Ⅰ-A6 包括PD6186、新陆早38 号、中R1029、豫棉19、辽棉15 号、LA887/宿Bt F9、A41772BBt F7、中资9103、运93 抗393,丰产耐旱品种。Ⅰ-A7 包括时09、中1816、抗黄萎164、中R014121。Ⅰ-A8 包括Ari971、中RI015、CZA(70)33、中AR40772、TM1-IPR、中远9115、L142-9、十五创新种质,耐旱、耐盐、抗虫。Ⅰ-A9 包括6 份种质,鲁棉2 号、徐州514、A41772152 F6、14214612 F5、SGK 石选321、辽棉-5 号。

表4 相似系数统计分析
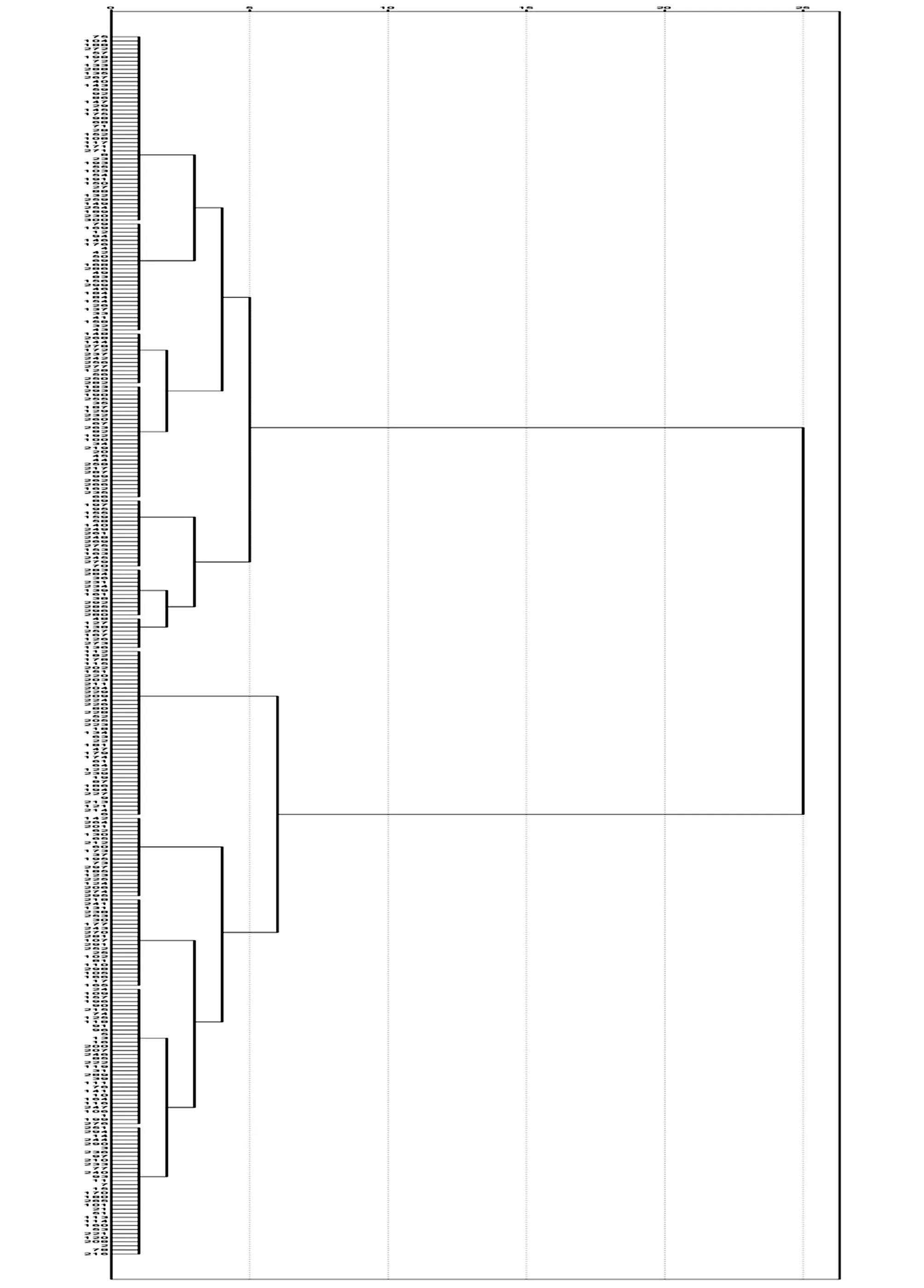
图1 基于表型性状的聚类图
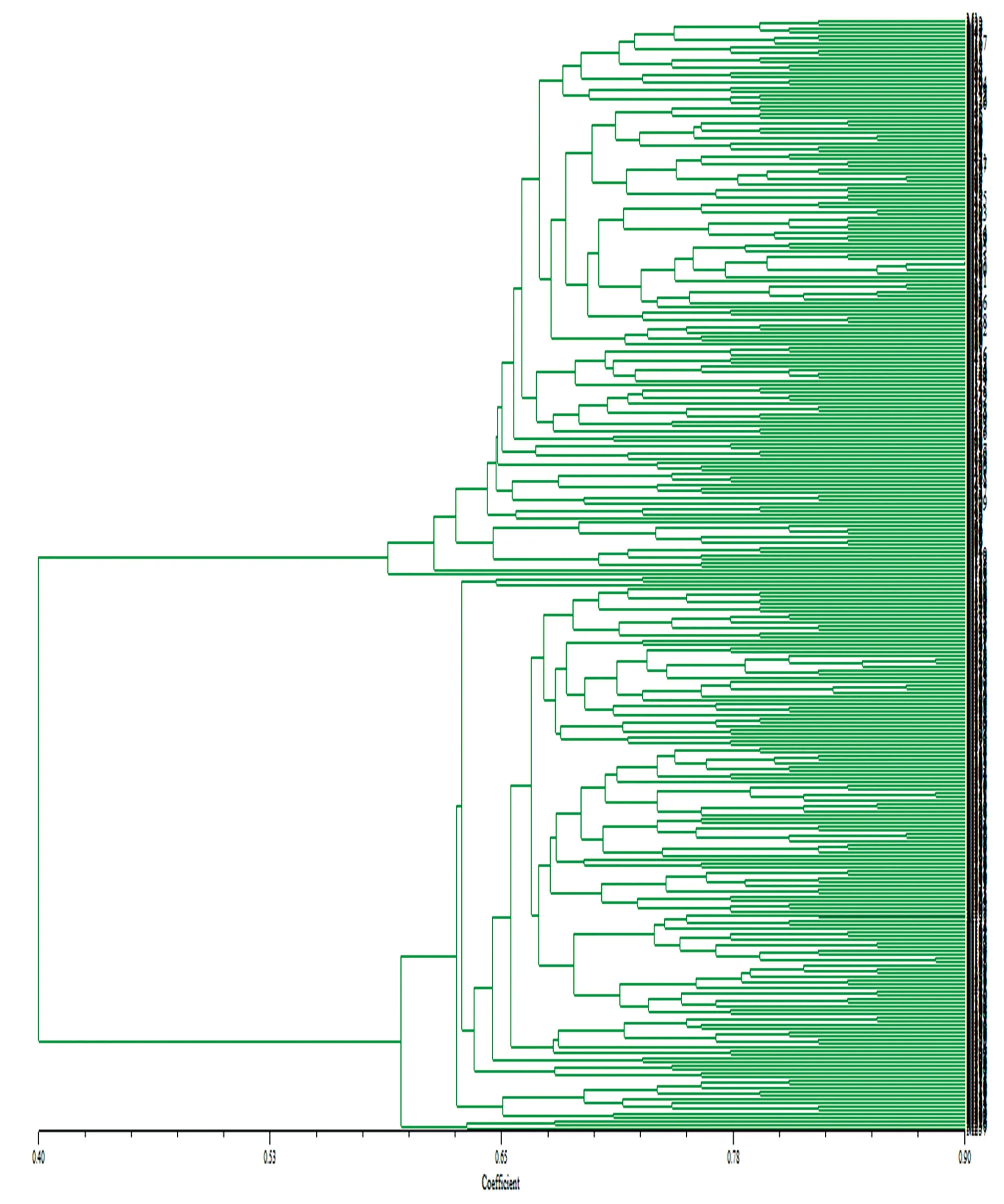
图2 基于SSR 的聚类图
类群Ⅱ包括150 个品种,阈值为0.594 时又被分为Ⅱ-1、Ⅱ-2 两大类。Ⅱ-1 在遗传相似系数为0.632 时被分为两类,Ⅱ-A 包括136 个品种,遗传相似系数变幅为0.635~0.891;Ⅱ-B 包括11 个品种,遗传相似系数变幅为0.655~0.828。Ⅱ-2 包含3 个品种,分别为838、川棉-239、纳上区大花,遗传相似系数为0.638~0.683。
Ⅱ-A 又被分为8 个亚类。Ⅱ-A1 包括11 个品种,塔什干6 号、湘163、川R128、苏BR6202Bt、J02-247、石抗278、澳B 资10、鄂沙28、泗长2 系、CS50、苏农6 号,大部分为创新种质。Ⅱ-A2 包括豫284、中远911、中G5。Ⅱ-A3 包括奎屯80-2056W、AcalaSJ-1-9、34 系、Miscot7803-52。Ⅱ-A4 包括永济1 号、中31-204。Ⅱ-A5 包括229、285、296、298、186、277、279、292、248、234、297 等11 个品种,无鲜明特点。Ⅱ-A6 包括181、177、172、164 等28 个品种,大部分为创新种质,丰产,来源较近。Ⅱ-A7 包括158、214、181、202、231 等45 个品种,大体上均抗虫,丰产,抗黄萎病。Ⅱ-A8 包括180、281、190、212、154 等43 个品种,株高比较高,主要是一些具有不同特殊品质的高蛋白、高酚等优质品种。
3 讨论
前人大量研究表明,聚类分析结果与该试验所选用的引物本身及引物数量有很大关系,考虑成本和效率的前提下,引物数量越多越好[15-17]。为了提高试验的准确性,我们尽可能多地使用SSR 引物。筛选出的38 对多态性引物均已定位在棉花染色体上,如果这些引物能够覆盖全部连锁群,试验结果将会更加理想。
本试验所筛选出的多态性引物数量比较少,多态性比率不高,很可能与采用随机选取法选取16 个品种进行引物初筛有关,应选用地理来源或表型性状差异较大的品种进行初筛。
表型性状和SSR 分子标记分析均被应用于棉花种质资源遗传多样性的研究,但2 种类型的分析结果各有侧重。表型性状调查结果是对观测性状的直接表现,而分子标记则揭示了个体在基因组水平上的基因结构之间的部分差异,只有将不同类型的数据结合起来使用,才能比较全面准确地阐明品种之间的遗传关系[18-21]。
本试验就将这2 种分析方式结合,通过分析可知,这2 种聚类结果既有一致的地方,也存在差异。例如,在2 种聚类结果中,总的大类聚类结果是一致的,43、26 和33 号等材料在表型聚类和分子标记聚类中被聚在同一类,类似的在2 种分类结果中得到相同聚类结果的材料所占比例很高。但是有些试验材料在2 种聚类方法中却表现出较大的差异,如1、109 号材料在形态性状聚类中的亲缘关系极近,但在SSR 分子标记聚类中却被聚在了同一大类的2 个亚类。
总之,2 种方法在大的类群划分上表现出比较好的一致性,但在类群内的划分出现了较大差异。这表明所选试验材料之间的遗传差异大小的变化,很可能会导致用遗传相似系数与遗传距离来确定的品种之间的遗传差异的变化。分子标记从DNA 分子水平揭示的多样性信息量大,更能比较真实地反映多态性的水平,本试验表明了分子标记用于检测遗传多样性的可靠性。在育种选种时,可以依据品种之间的遗传相似系数的高低来选择。但是,在实际中很难评定哪种聚类方法更好,要根据研究目的而定,对于以遗传多样性为基础的核心种质的构建来说,既要考虑基因型,又要考虑表现型,同时还要兼顾一些特异种质资源,所以要多层面、多角度地分析研究,多种方法结合使用并相互印证,才能比较全面地了解种质之间的亲缘关系。
综上,表型和SSR 聚类分析表明本实验室保存的300 份棉花种质资源遗传多样性指数较高,表现出较丰富的遗传多样性,为培育高产稳产早熟的棉花新品种提供了理论依据。
猜你喜欢
今日农业(2022年14期)2022-09-15
今日农业(2022年13期)2022-09-15
世界科学技术-中医药现代化(2022年3期)2022-08-22
昆明医科大学学报(2021年3期)2021-07-22
四川蚕业(2020年4期)2020-02-10
中国麻业科学(2018年6期)2018-04-09
现代园艺(2017年21期)2018-01-03
中国康复理论与实践(2015年10期)2015-12-24
中央民族大学学报(自然科学版)(2015年1期)2015-06-11
医学研究杂志(2015年5期)2015-06-10
