
基于iTRAQ技术的甘蔗受黑穗病菌侵染蛋白组分析
2019-12-25 10:53宋奇琦PratikshaSINGHRajeshKumarSINGH宋修鹏李海碧农友业杨丽涛李杨瑞
作物学报 2019年1期
宋奇琦 Pratiksha SINGH Rajesh Kumar SINGH 宋修鹏 李海碧农友业 杨丽涛,,* 李杨瑞,,*
基于iTRAQ技术的甘蔗受黑穗病菌侵染蛋白组分析
宋奇琦1,**Pratiksha SINGH2,**Rajesh Kumar SINGH2宋修鹏1李海碧1农友业2杨丽涛1,2,*李杨瑞1,2,*
1中国农业科学院甘蔗研究中心/ 广西壮族自治区农业科学院/ 农业部广西甘蔗生物技术与遗传改良重点实验室/ 广西甘蔗遗传改良重点实验室, 广西南宁 530007;2广西大学农学院/ 亚热带农业生物保护与利用国家重点实验室, 广西南宁 530005
黑穗病已成为影响甘蔗产量和含糖量的重要病害。为从蛋白质水平探讨甘蔗应答黑穗病菌的分子机制, 本实验选用抗黑穗病品种桂糖29号和感黑穗病品种崖城71-374, 处理组用浸渍法接种黑穗病菌, 对照组用无菌水模拟接菌, 在接种180 d后采取甘蔗叶片, 使用iTRAQ技术对蛋白质组进行研究。结果显示, 桂糖29号中有定量信息蛋白1429个, 差异表达蛋白290个, 其中上调表达蛋白153个, 下调表达蛋白137个; 崖城71-374中有定量信息蛋白1576个, 差异表达蛋白125个, 其中上调表达蛋白55个, 下调表达蛋白70个。抗病品种桂糖29号中差异表达蛋白数多于感病品种崖城71-374, 且桂糖29号在KEGG富集到的代谢通路也更多, 可能被侵染后抗病品种的免疫调节机制更为复杂, 涉及的调控通路网更广。经对光合作用、抗氧化系统、钙信号、苯丙烷类代谢、激素相关差异表达蛋白及共有差异表达蛋白分析, 发现光合作用通路、ROS、ABA、钙信号通路相关蛋白在2个品种中多为上调表达, 且桂糖29号的上调表达蛋白数多于崖城71-374, 可能参与甘蔗后期对黑穗病的应答。植物抗病是一个复杂的过程, 需要多种功能与途径参与调控。在本实验中没有发现苯丙烷类代谢途径及一些酶(谷胱甘肽过氧化物酶、抗坏血酸过氧化物酶、超氧化物歧化酶、谷胱甘肽硫转移酶、过氧化氢酶)、激素(生长素、乙烯、赤霉素)参与甘蔗的抗病过程, 可能与采样时间有关。
甘蔗; 黑穗病; iTRAQ; 蛋白质组
甘蔗是重要的农作物, 种植在包括中国在内的90多个国家。黑穗病已经成为全球性的甘蔗病害, 且发病率逐年提高。如果感染严重, 可造成甘蔗20%~50%的产量损失, 糖含量也会大量减少[1-2]。
前人关于黑穗病的研究大部分集中在生理生化方面。Lloyd等[3]采用越过芽麟障碍的针刺接种方法, 将病原菌注入一些甘蔗品种芽体分生组织区后, 发现有些甘蔗品种仍不发病, 表明甘蔗体内产生了抑制病原菌入侵繁殖的物质, 同时也揭示了甘蔗品种生理生化抗性的存在。Singh等[4]观察到2个易感黑穗病甘蔗品种的叶片、芽、顶端分生组织、侧枝以及茎中甘蔗汁的抗坏血酸含量增加, 推测这可能是病原菌产生抗坏血酸加速酶或宿主-病原菌相互作用所致。Santiago等[5]研究由黑穗病诱导植物酚类物质含量的变化, 导致细胞壁木质化加厚, 可作为对病原菌入侵的机械防御, 推测木质素在机械组织中的沉积会增加甘蔗的生化和形态学抗性反应。莫凤莲等[6]利用透射电子显微镜观察甘蔗感黑穗病和蔗芽的超微结构变化, 发现其细胞变形严重甚至空泡化, 细胞核、核仁、染色质和线粒体等均发生改变, 同时研究甘蔗接种黑穗病菌后内源激素含量变化, 甘蔗叶片中生长素(IAA)、赤霉素(GA)、脱落酸(ABA)、水杨酸(SA)含量及IAA/ABA值的变化与甘蔗抗性密切相关。苏亚春等[7]研究表明, 甘蔗抗性品种崖城05-179在感染黑穗病初期几丁质酶、β-1,3-葡聚糖酶、过氧化氢酶活性均高于感病品种柳城03-182, 这些酶活性与甘蔗黑穗病抗性有一定联系。Leila等[8]研究发现, 抗性甘蔗品种中过氧化氢(H2O2)积累与甘蔗鞭黑粉菌冬孢子在植物体内萌发和附着胞形成阶段一致, 而感病品种只有少量H2O2的积累, 但2个品种均没有超氧阴离子自由基(O2–)的累积, 同时通过对丙二醛(MDA)、过氧化氢酶(CAT)、谷胱甘肽-S转移酶(GST)活性及蛋白质组研究, 推测H2O2的积累可能是与抗坏血酸、谷胱甘肽和硫氧还蛋白相关的多重系统来影响信号传导。
随着先进生物技术手段的出现, 人们对甘蔗-病原菌互作有了更深入的了解。最近的分子生物学研究涉及分子标记技术、转录组学和蛋白质组学等。宋修鹏[9]成功构建了甘蔗幼苗应答黑穗病侵染的正向SSH文库, 并利用同源克隆和RACE技术得到苯丙氨酸解氨酶(PAL)、S-腺苷甲硫氨酸合成酶(SAM)、木葡聚糖内糖基转移水解酶(XTH)、Nudix水解酶(NUDT)基因全长。Barnabas等[10]通过双向电泳(2-DE)联合MALDI-TOF/TOF-MS技术鉴定感黑穗病40 d后甘蔗差异蛋白, 鉴定出53个蛋白质与防御、胁迫、代谢、蛋白质折叠、能量和细胞分裂有关。Su等[11]通过蛋白质组与转录组关联分析发现两者的相关性较差, 与差异基因表达趋势相同的蛋白大多与植物抗逆相关, iTRAQ分析结果显示, 乙烯、赤霉素、苯丙烷类代谢参与甘蔗抗黑穗病过程, 而钙信号通路、活性氧通路、脱落酸没有参与应答过程。Que等[12]利用2-DE技术分析甘蔗品种NCo376和Ya71-374感黑穗病后蛋白质的表达, 生物信息学分析表明, 20 个差异蛋白与光合作用、信号转导等功能有关。
同位素相对和绝对定量标记(isobaric tags for relative and absolute quantitation, iTRAQ)是2004年美国ABI公司研发的一种新型定量蛋白质组学技术, 该技术具有分离能力强、定性分析结果可靠、灵敏度高等优点[13]。唐成等[14]基于iTRAQ技术对稻瘟病侵染水稻幼苗叶片后差异表达蛋白分析, 表明53个差异蛋白涉及氧化还原平衡、防御、信号传导、糖和能量代谢、氨基酸代谢、光合作用及蛋白质代谢等途径。Marsh等[15]利用iTRAQ技术分析葡萄感染白粉病后叶片在不同时间点的差异表达蛋白, 鉴定到63个差异蛋白与光合作用、代谢、病害防御、蛋白质合成等相关。目前对甘蔗黑穗病的研究大部分在感病初期, 本实验利用iTRAQ技术分析甘蔗与黑穗病长期互作后差异表达蛋白, 探寻与抗黑穗病相关的功能及代谢途径, 以期为甘蔗抗黑穗病分子机制的研究提供进一步的理论依据。
1 材料与方法
1.1 材料
选用甘蔗品种桂糖29号(GT29)和崖城71-374 (Yacheng 71-374), 由广西农业科学院甘蔗研究所提供。其中, GT29经多年试验, 其新植和宿根均表现为高抗黑穗病[16]; Yacheng 71-374经鉴定为高感黑穗病品种[17]。黑穗病菌混合冬孢子收集于广西大学试验田甘蔗品种桂糖11号、桂选B9、新台糖22号、拔地拉。
1.2 方法
1.2.1 样品采集 2016年4月至11月在广西大学甘蔗智能温室大棚, 将蔗种砍成单芽段, 水浴锅中50℃浸种2 h, 之后进行沙培育苗。1个月后, 分别挑选每个品种60株长势一致的幼苗, 用无菌水清洗后分成两等份。一组经黑穗病菌孢子浓度为5106个孢子 mL–1悬浮液浸泡2 h作为处理组, 另一组则用无菌水浸泡2 h作为对照组, 将幼苗移栽到桶里, 置于甘蔗智能温室。甘蔗通常在病菌侵染6~7个月黑穗长度达到顶峰[18], 所以选取感染180 d后时间点取样。采用浸渍法接菌最接近黑穗病菌侵染甘蔗的自然状态, 在侵染180 d后, 感病甘蔗比对照植株矮小, 叶片细长, 但叶片颜色肉眼观察差异不明显。均取2个品种甘蔗发病植株+1叶, 去除叶脉, 采集的样品经蒸馏水清洗后擦干, 用液氮冷冻, 保存于-80°C。2个品种均有2个生物学重复。
1.2.2 PCR检测 使用植物DNA提取试剂盒(康为世纪, 北京)从GT29和Yacheng 71-374感病及对照植株中提取基因组DNA; 通过HP Fungal DNA Kit (Omega, 中国宁波)试剂盒提取黑穗病冬孢子DNA作为阳性对照, 无菌水为阴性对照。用黑穗病菌检测引物bE4 (5¢-CGCTCTGGTTCATCAACG-3')和bE8 (5¢-TGCTGTCGATGGAAGGTGT-3¢)检测甘蔗是否感染黑穗病菌。用引物bE4/bE8能扩增出黑穗病菌基因组序列上的一段特异序列, 长度为459 bp。
1.2.3 蛋白样品制备 提取蛋白参考林丽等[19]酚抽提法, 用裂解液溶解蛋白, 加入5倍体积预冷的丙酮, 于-20°C沉淀过夜, 离心得到蛋白沉淀, 晾干10 min左右, 用iTRAQ定量试剂盒中dissolution buffer溶解。
1.2.4 蛋白质样品定量与酶解 按照Qubit Protein Assay Kit试剂盒说明书进行蛋白质定量, 根据定量结果, 从每个样品精确取出100 μg蛋白, 加入iTRAQ定量试剂盒中1 μL 2% SDS溶液使蛋白质变性, 然后再加入2 μL还原试剂, 涡旋混合, 于60°C孵育1 h后加入1 μL半胱氨酸封闭试剂反应30 min, 加入1 μg μL–1胰蛋白酶, 胰蛋白酶与蛋白质的质量比为1∶20, 37°C酶解12~16 h。
1.2.5 用iTRAQ试剂标记 取出iTRAQ标记, 室温平衡, 短暂离心至管底。每管加入70 µL 乙醇, 震荡混匀, 短暂离心。将每管标记试剂分别转移至样品管中, 4个样品分别用不同的同位素标记, 即用iTRAQ 8 Plex 114标记GT29对照, 115标记GT29处理, 116标记Yacheng 71-374对照, 117标记Yacheng 71-374处理。震荡混匀, 测定溶液的pH值, 保证pH=7.8~8.5, 且保证溶液的有机相比例≥70%, 室温孵育1 h, 然后加入一定量的超纯水(约100 µL)终止标记反应。混合各管样品, 震荡混匀, 短暂离心。使用C18小柱去除杂质, 真空浓缩抽干备用。
1.2.6 用高pH值C18反相色谱柱进行第一维多肽分离 上述标记后, 将样品复溶于流动相A, 使用C18反相色谱柱XBridge Peptide BEHC18 (4.6 mm×250.0 mm)进行分离收集。流动相A为98%水, 2%乙腈, pH 10.0。流动相B为98%乙腈, 2%水, pH值为10.0。流速0.5 mL min–1。梯度0~8 min, 100% A; 8~48 min, 100% A~40% B; 48~53 min, 40%~100% B; 53~63 min, 100% B; 63~65 min, 100% B~100% A; 65~75 min, 100% A。检测波长为214 nm。从8 min开始收集馏分, 每2 min收集1管, 然后将首尾收集的馏分适当合并, 最后得到10个馏分。
1.2.7 纳升液相结合LTQ-Orbitrap质谱分析 将第一维分离后的馏分, 通过脱盐处理、真空抽干, 复溶于流动相A, 以便进行后续的LC-MS/MS 分析。流动相A为2% ACN+0.1% HCOOH。流动相B为98% ACN+0.1% HCOOH。洗脱梯度: 0~5 min, 2%~10% B; 5~50 min, 10%~35% B; 50~55 min, 35%~98% B; 55~65 min, 98% B。流速为250 nL min–1。色谱柱为C18反相毛细管柱。
质谱模式为ESI正离子模式, 一级质谱扫描分辨率为60,000; 扫描范围为m/z = 350~1800; 二级质谱扫描分辨率为15,000, 用HCD 碰撞模式完成, 采用数据依赖型的自动化采集模式。电喷雾电压为2.8 kV。
1.2.8 质谱数据的蛋白质数据库搜寻鉴定 质谱采集到的原始数据采用Proteome Discovery软件的SEQUEST搜索器进行搜索和匹配, 采用本课题组测定的转录组翻译的蛋白数据库。搜索参数设置如下, 消化酶胰蛋白酶(trypsin)作为消化酶, 选择酶解最大2个漏切位点, 固定修饰为甲硫氨酸(methionine)的氧化, 可变修饰为烷基化(carbamidomethyl C)以及iTRAQ对N端和氨基酸K、Y位点的修饰。
1.2.9 生物信息学分析 对鉴定到的蛋白进行GO注释, 并进一步对差异表达蛋白进行GO富集、KEGG富集分析。分别对每个甘蔗品种的2个样本进行重复性分析。2个样本的ratio取平均值, 并通过2次结果进行-test检验, 计算-value值, 当-value<0.05, ratio>1.2或ratio<0.83视为差异表达蛋白。
2 结果与分析
2.1 PCR检测结果
琼脂糖凝胶电泳表明, GT29和Yacheng 71-374接种黑穗病的植株与阳性对照均可扩增出459 bp左右的条带, 对照植株则无此条带(图1)。扩增的PCR片段序列与GenBank中公布的bEast交配型基因序列相同, 证实处理组甘蔗已感染黑穗病菌, 而对照组没有被感染(GenBank登录号为KY575283~KY575284)。
2.2 鉴定的蛋白质信息
成功鉴定的有定量信息的蛋白共有1831个, 其中共有蛋白1174个。Yacheng 71-374中鉴定到的蛋白数比GT29中多147个。GT29鉴定到的差异表达蛋白有290个, 其中上调表达蛋白有153个, 下调表达蛋白有137个。Yacheng 71-374中鉴定到的差异表达蛋白有125个, 其中上调表达蛋白有55个, 下调表达蛋白有70个。抗病品种GT29中差异表达蛋白数多于感病品种Yacheng 71-374 (表1)。
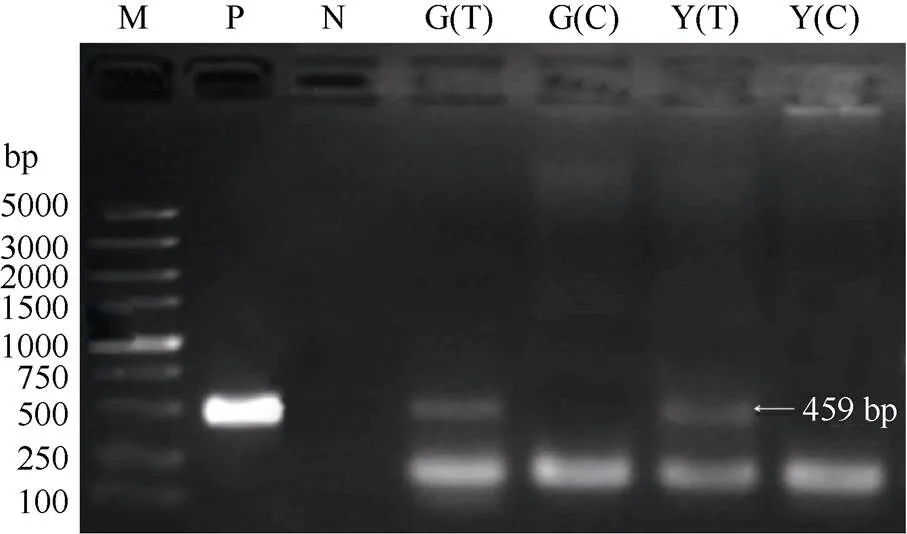
图1 对照和接种黑穗病菌甘蔗DNA的PCR扩增产物电泳图
M: molecular size marker (100~5000 bp); P: 黑穗病孢子DNA正对照; N: 无菌水负对照; G(T): GT29处理; G(C): GT29 对照; Y(T): Yacheng 71-374处理; Y(C): Yacheng 71-374对照。
M: molecular size marker (100-5000 bp); P:DNA as positive; N: sterile water as negative; G(T): GT29 treatment; G(C): GT29 control; Y(T): Yacheng 71-374 treatment; Y(C): Yacheng 71-374 control.

表1 2个甘蔗品种中鉴定到的肽段和蛋白数
GO注释包含生物过程(biological process)、分子功能(molecular function)、细胞组分(cellular component)三大分支。对总蛋白进行GO注释, 鉴定到的蛋白在生物过程中占比例最大条目为代谢过程(metabolic process)(62%)、细胞过程(cellular process)(43%)、单一生物过程(single-organism process)(35%), 在分子功能中催化(catalytic)(55%)和结合(binding)(48%)占多数, 而细胞组分中主要条目为细胞(cell)(25%)和细胞部分(cell part)(25%)(图2)。
2.3 差异表达蛋白GO富集分析
GT29差异表达蛋白GO富集分析结果如附表1所示, 在生物过程中, 极显著富集(<0.01)条目有光合作用-光反应(photosynthesis-light reaction)、光合作用-光捕获(photosynthesis-light harvesting)、光合作用(photosynthesis); 分子功能中主要富集条目为蛋氨酸腺苷转移酶活性(methionine adenosyl transferase activity)、酶抑制剂活性(enzyme inhibitor activity); 细胞组分中极显著富集(<0.01)条目为膜(membrane)。

图2 GO注释分析
Yacheng 71-374差异表达蛋白富集结果如附表2所示。生物过程中极显著富集(<0.01)条目有氧化应激反应(response to oxidative stress)、代谢调节过程(regulation of metabolic process); 而分子功能中极显著富集(<0.01)条目有7条, 分别为儿茶酚氧化酶活性(catechol oxidase activity)、氨裂解酶活性(ammonia-lyase activity)、氧化还原酶活性(oxidoreductase activity, acting on diphenols and related substances as donors, oxygen as acceptor)、碳酸脱水酶活性(carbonate dehydratase activity)、氧化还原酶活性(oxidoreductase activity acting on peroxide as acceptor)、过氧化物酶活性(peroxidase activity)、裂解酶活性(lyase activity); 细胞组分中主要富集到光合体系(photosystem)条目中。
2.4 差异表达蛋白KEGG富集分析
通过kyoto encyclopedia of genes and genomes (KEGG)数据库对差异蛋白进行富集分析。GT29差异蛋白显著富集(<0.05)到8条通路中, 分别为核糖体(ribosome)、光合作用(photosynthesis)、类胡萝卜素生物合成(carotenoid biosynthesis)、植物-病原菌互作(plant-pathogen interaction)、抗原的加工提呈(antigen processing and presentation)、双组分系统(two-component system)、光合作用-天线蛋白(photosynthesis-antenna proteins)、吞噬体(phagosome)。其中, 极显著富集(<0.01)通路为光合作用(=0.0067)、核糖体(=0.0063)。
Yacheng 71-374差异蛋白显著富集(<0.05)到3条通路中, 分别为光合作用(photosynthesis)、苯丙烷类生物合成(phenylpropanoid biosynthesis)、次生代谢产物生物合成(biosynthesis of secondary metabolites), 其中极显著富集(<0.01)通路为光合作用(= 0.0014)、苯丙烷类生物合成(=0.0082)。
2.5 光合作用相关差异表达蛋白
GO富集和KEGG富集结果显示, 2个甘蔗品种均在光合作用代谢过程差异蛋白最多、差异最显著, 因此, 我们重点关注了与此相关的差异表达蛋白。光合作用4个多亚基蛋白复合体分别为光系统I (PS I)、光系统II (PSII)、ATP合成酶(ATPase)及细胞色素b6f复合体(Cytb6f)[20]。PS I由天线系统、反应中心、外周蛋白、内在蛋白构成, PS II主要由外周天线、内周天线、反应中心、放氧复合体和辅助亚基构成[21]。由表2可知, GT29鉴定到28个相关差异蛋白, 26个上调表达, 2个下调表达; Yacheng 71-374鉴定到16个相关差异蛋白, 12个上调表达, 4个下调表达。PSI反应中心亚基在GT29中有4个(c54318_ g1;orf1、c69501_g1;orf1、c56023_g1;orf1、c56941_g1; orf1)均为上调表达, 在Yacheng 71-374中有5个(c54318_g1;orf1、c48673_g1;orf1、c43081_g1;orf1、c129327_g1;orf1、c55121_g1;orf1)同样都上调表达; 铁氧还蛋白[2Fe-2S]为PSI反应中心电子受体[22], 在GT29下调表达, Yacheng 71-374无差异; PSI组装蛋白ycf4在GT29上调表达, Yacheng 71-374下调表达; 叶绿素结合蛋白由Lhca基因家族编码, 为PSI天线系统[23], 在GT29有11个差异表达, 其中仅有1个下调表达, 在Yacheng 71-374有4个差异表达, 其中1个下调表达。PSII反应中心相关蛋白[21]在GT29中PSII蛋白D1、PSII蛋白D2、细胞色素b559亚基α均上调表达, 在Yacheng 71-374仅细胞色素b559亚基α (c54219_g1;orf2)差异下调; 内周天线蛋白PSII CP43脱辅基蛋白(c77580_g1;orf1)在GT29上调表达, Yacheng 71-374中无差异; 放氧增强蛋白是PSII的放氧复合体的一部分[24], 结果显示, GT29中有1个上调表达, Yacheng 71-374有2个上调表达; PSII PsbR、PSII 47 kDa蛋白在GT29上调表达, 在Yacheng 71-374中无差异。细胞色素b6f复合体连接PSI和PSII中电子传递, 同时促进ATP合成。GT29中细胞色素b6、细胞色素f上调表达, 而在Yacheng 71-374中均无差异。ATP合成酶在GT29中一个上调表达一个下调表达, 在Yacheng 71-374中均无差异。还有叶绿体Ptr-ToxA结合蛋白在GT29上调表达, 铜蛋白在Yacheng 71-374上调表达, 16 kDa膜蛋白在GT29上调表达, 而在Yacheng 71-374下调表达。结果表明, 相关差异蛋白参与光合作用的多个方面, 可能在2个甘蔗品种抗黑穗病过程均起重要作用, 在抗病品种GT29中表现更加突出。
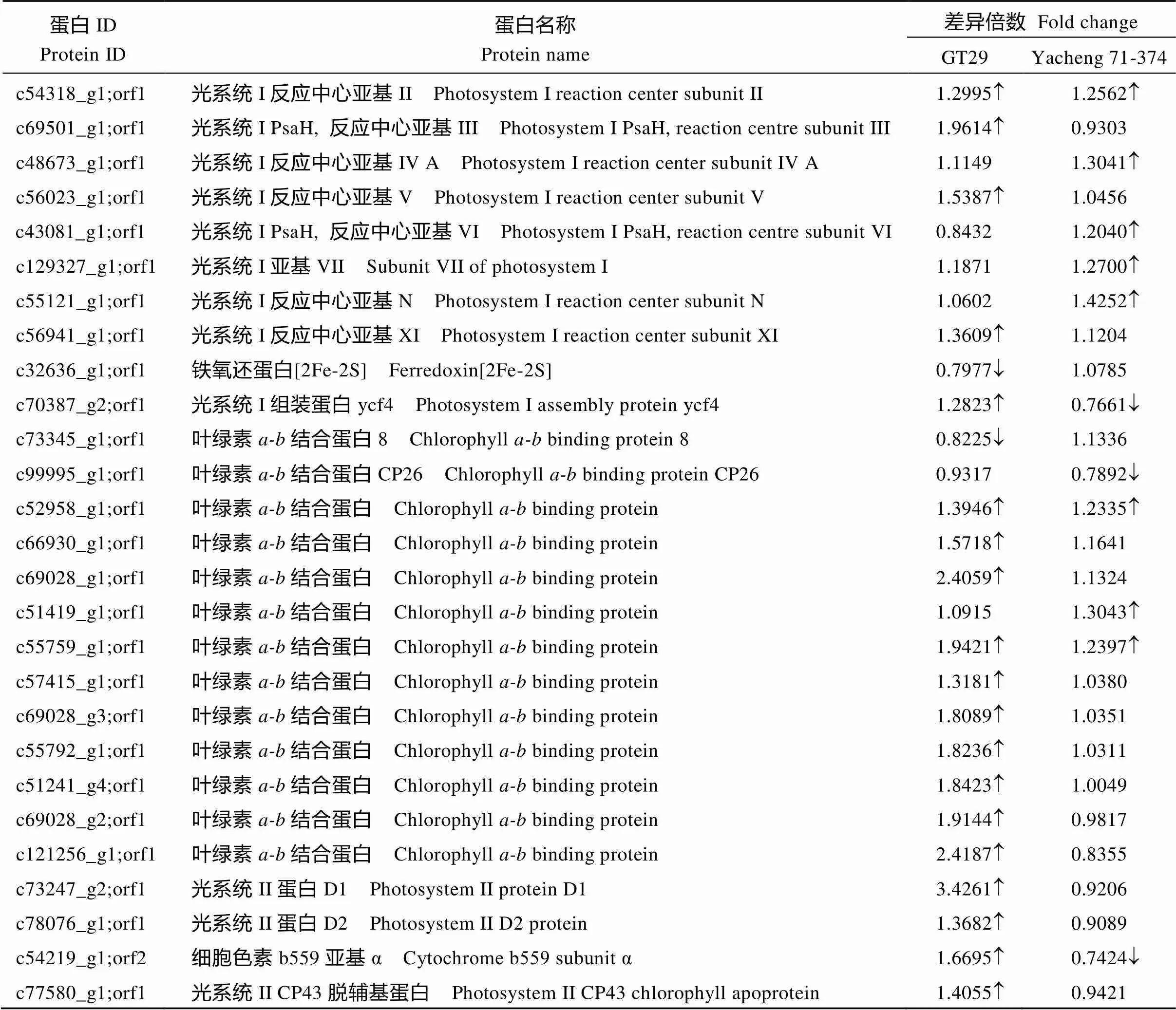
表2 光合作用相关差异表达蛋白
(续表2)

蛋白IDProtein ID蛋白名称Protein name差异倍数 Fold change GT29Yacheng 71-374 c126236_g1;orf1放氧增强蛋白1 Oxygen-evolving enhancer protein 11.00641.2372 c59540_g1;orf1放氧增强蛋白3 Oxygen-evolving enhancer protein 30.87541.8631 c100226_g1;orf1放氧增强蛋白 Oxygen-evolving enhancer protein1.53060.9906 c55903_g1;orf1叶绿体放氧增强蛋白1 (部分) Chloroplast oxygen-evolving enhancer protein 1 (partial)1.04561.2517 c51741_g1;orf1光系统II PsbR Photosystem II PsbR1.90371.0547 c22663_g1;orf1光系统II 47kDa蛋白 Photosystem II 47 kDa protein1.72950.9404 c56158_g2;orf1细胞色素b6 (部分) Cytochrome b6 (partial)1.47861.0816 c72546_g2;orf1细胞色素f Cytochrome f1.25400.9964 c67723_g1;orf1ATP合成酶CF1α亚基 ATP synthase CF1 α subunit1.38371.0172 c57360_g1;orf2ATP合成酶 ATP synthase0.6735¯1.0127 c61429_g2;orf1叶绿体Ptr-ToxA结合蛋白 Chloroplast Ptr ToxA-binding protein1.41160.9843 c54911_g1;orf116 kDa膜蛋白 16 kDa membrane protein1.63640.7557¯ c47557_g1;orf1铜蛋白 Blue (type 1) Copper protein Blue (type 1)1.11701.5826
为上调表达,¯为下调表达。is up-regulated and¯is down-regulated.
2.6 抗氧化相关差异表达蛋白
植物在受到胁迫时会产生活性氧(ROS), 而过量的ROS会氧化蛋白质、脂质和核酸, 从而导致细胞结构的损伤[25]。鉴定到的相关酶包括过氧化物酶(POD)、谷胱甘肽过氧化物酶(GPX)、抗坏血酸过氧化物酶(APX)、超氧化物歧化酶(SOD)、谷胱甘肽硫转移酶(GST)和过氧化氢酶(CAT)。
由表3可知, 在GT29中鉴定到15个POD差异表达蛋白, 其中11个上调表达; Yacheng 71-374鉴定到13个差异表达蛋白, 其中10个上调表达, 3个下调表达。在GT29鉴定到2个GPX差异表达蛋白均下调表达, 而在Yacheng 71-374中无差异表达。在GT29鉴定到4个APX差异表达蛋白均下调表达; 在Yacheng 71-374中鉴定到1个差异下调表达蛋白。在GT29鉴定到3个SOD差异表达蛋白均下调表达; 在Yacheng 71-374中鉴定到1个差异上调表达蛋白。在GT29鉴定到6个GST差异表达蛋白其中1个上调表达, 5个上调表达; 而在Yacheng 71-374中鉴定到5个差异表达蛋白均为上调表达。在GT29鉴定到3个CAT差异表达蛋白, 其中1个上调表达, 2个下调表达; 在Yacheng 71-374中鉴定到1个差异下调表达蛋白。2个品种的POD均大部分为上调表达, 抗性品种GT29稍多一些,而GPX、APX、SOD、GST、CAT在GT29中大都为下调表达, Yacheng 71-374中表现为上调或无差异, 结果表明, 甘蔗在受黑穗病菌胁迫时会诱导抗氧化蛋白来参与应答, 可能主要为POD。
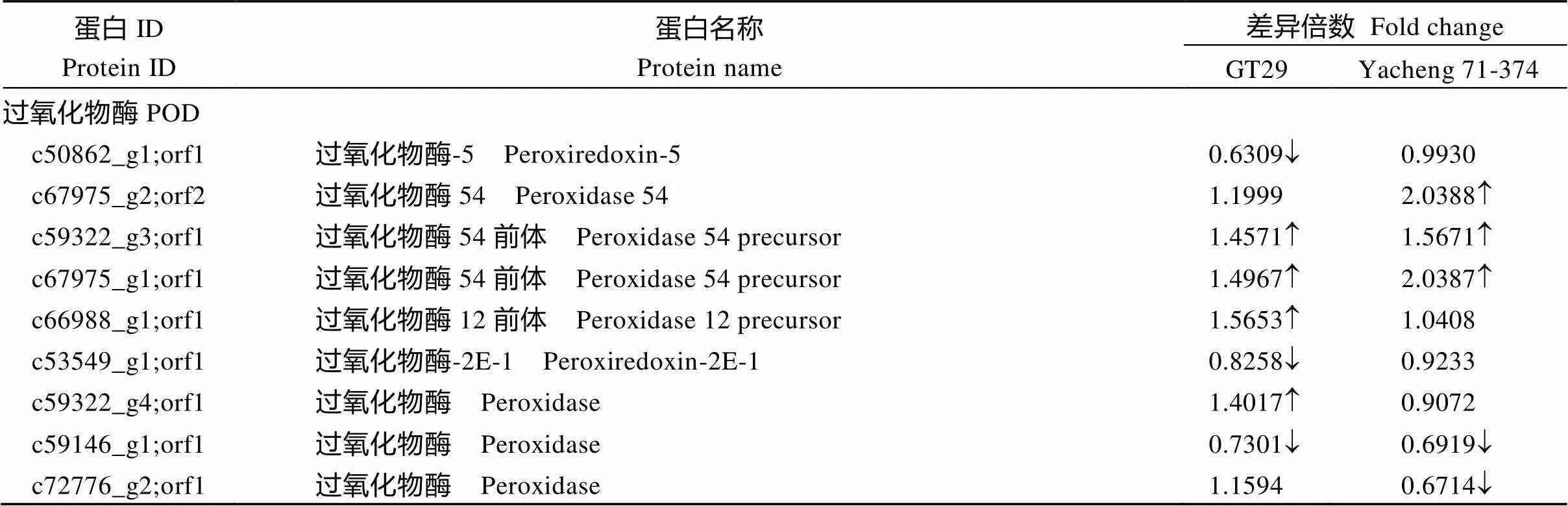
表3 抗氧化相关差异表达蛋白
(续表3)
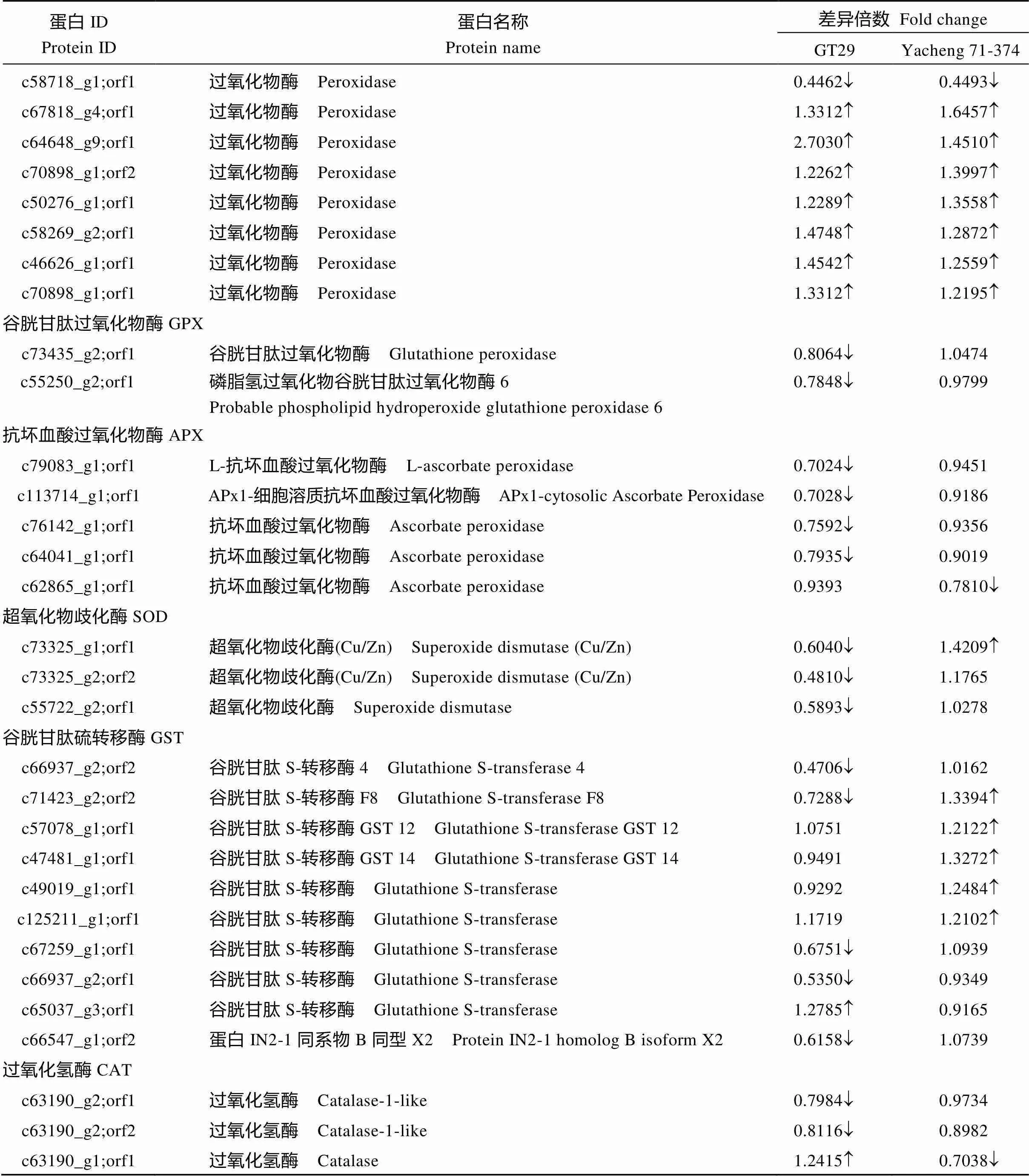
蛋白IDProtein ID蛋白名称Protein name差异倍数 Fold change GT29Yacheng 71-374 c58718_g1;orf1过氧化物酶 Peroxidase0.4462¯0.4493¯ c67818_g4;orf1过氧化物酶 Peroxidase1.33121.6457 c64648_g9;orf1过氧化物酶 Peroxidase2.70301.4510 c70898_g1;orf2过氧化物酶 Peroxidase1.22621.3997 c50276_g1;orf1过氧化物酶 Peroxidase1.22891.3558 c58269_g2;orf1过氧化物酶 Peroxidase1.47481.2872 c46626_g1;orf1过氧化物酶 Peroxidase1.45421.2559 c70898_g1;orf1过氧化物酶 Peroxidase1.33121.2195 谷胱甘肽过氧化物酶GPX c73435_g2;orf1谷胱甘肽过氧化物酶 Glutathione peroxidase0.8064¯1.0474 c55250_g2;orf1磷脂氢过氧化物谷胱甘肽过氧化物酶6Probable phospholipid hydroperoxide glutathione peroxidase 60.7848¯0.9799 抗坏血酸过氧化物酶APX c79083_g1;orf1L-抗坏血酸过氧化物酶 L-ascorbate peroxidase0.7024¯0.9451 c113714_g1;orf1APx1-细胞溶质抗坏血酸过氧化物酶 APx1-cytosolic Ascorbate Peroxidase0.7028¯0.9186 c76142_g1;orf1抗坏血酸过氧化物酶 Ascorbate peroxidase0.7592¯0.9356 c64041_g1;orf1抗坏血酸过氧化物酶 Ascorbate peroxidase0.7935¯0.9019 c62865_g1;orf1抗坏血酸过氧化物酶 Ascorbate peroxidase0.93930.7810¯ 超氧化物歧化酶SOD c73325_g1;orf1超氧化物歧化酶(Cu/Zn) Superoxide dismutase (Cu/Zn)0.6040¯1.4209 c73325_g2;orf2超氧化物歧化酶(Cu/Zn) Superoxide dismutase (Cu/Zn)0.4810¯1.1765 c55722_g2;orf1超氧化物歧化酶 Superoxide dismutase0.5893¯1.0278 谷胱甘肽硫转移酶GST c66937_g2;orf2谷胱甘肽S-转移酶4 Glutathione S-transferase 40.4706¯1.0162 c71423_g2;orf2谷胱甘肽S-转移酶F8 Glutathione S-transferase F80.7288¯1.3394 c57078_g1;orf1谷胱甘肽S-转移酶GST 12 Glutathione S-transferase GST 121.07511.2122 c47481_g1;orf1谷胱甘肽S-转移酶GST 14 Glutathione S-transferase GST 140.94911.3272 c49019_g1;orf1谷胱甘肽S-转移酶 Glutathione S-transferase0.92921.2484 c125211_g1;orf1谷胱甘肽S-转移酶 Glutathione S-transferase1.17191.2102 c67259_g1;orf1谷胱甘肽S-转移酶 Glutathione S-transferase0.6751¯1.0939 c66937_g2;orf1谷胱甘肽S-转移酶 Glutathione S-transferase0.5350¯0.9349 c65037_g3;orf1谷胱甘肽S-转移酶 Glutathione S-transferase1.27850.9165 c66547_g1;orf2蛋白IN2-1同系物B同型X2 Protein IN2-1 homolog B isoform X20.6158¯1.0739 过氧化氢酶CAT c63190_g2;orf1过氧化氢酶 Catalase-1-like0.7984¯0.9734 c63190_g2;orf2过氧化氢酶 Catalase-1-like0.8116¯0.8982 c63190_g1;orf1过氧化氢酶 Catalase1.24150.7038¯
为上调表达,¯为下调表达。is up-regulated and¯is down-regulated.
2.7 钙信号通路相关差异表达蛋白
钙信号是植物信号转导途径中重要的信号分子, 参与了大多细胞生理代谢过程[26]。在GT29中鉴定到2个差异表达钙调蛋白均下调表达, 1个钙依赖性蛋白激酶上调表达, 3个EF-手型钙结合蛋白中有1个上调表达, 2个下调表达。而在Yacheng 71-374中没有鉴定到任何相关差异表达蛋白。说明钙信号在一定程度上参与了甘蔗应答黑穗病过程, 但可能不是主要影响因素。
2.8 苯丙烷类代谢相关差异表达蛋白
在受到生物或非生物胁迫时, 苯丙烷类代谢途径是植物抗病反应次生代谢中重要的代谢途径之一[27]。本实验鉴定到相关差异蛋白中苯丙氨酸解氨酶是苯丙烷途径的关键酶和限速酶, 在GT29中有1个上调表达, 在Yacheng 71-374中有2个上调表达。3-脱氢奎尼酸合酶在苯丙氨酸生物合成途径中将4-磷酸赤藓糖催化生成分支酸[28], 其在GT29中无差异, 在Yacheng 71-374中上调表达。分支酸变位酶催化分支酸生成预苯酸[28], 在GT29中上调表达, 在Yacheng 71-374中无差异。DAHP合成酶受终产物酪氨酸和苯丙氨酸的协同反馈抑制[29], 在2个品种均下调表达。咖啡酰辅酶A-O-甲基转移酶是木质素生物合成的关键酶[30], 在2个品种均下调表达。表明苯丙烷类代谢途径可能也不是甘蔗抗黑穗病的关键途径。
2.9 激素相关差异表达蛋白
激素在甘蔗抗黑穗病中的作用已有研究[31]。与生长素(IAA)相关蛋白IAA-氨基酸水解酶ILR1、生长素抑制12.5 kDa蛋白在GT29中均下调表达, 在Yacheng 71-374中无差异。与乙烯(ETH)相关蛋白1-氨基环丙烷-1-羧酸酯氧化酶在GT29中无差异, 在Yacheng 71-374中下调表达。与脱落酸(ABA)相关蛋白, 在GT29中有3个蛋白磷酸酶2C、1个推定9-顺式-环氧类胡萝卜素双加氧酶均上调表达, 在Yacheng 71-374中有2个蛋白磷酸酶2C同样均上调表达。与赤霉素(GA)相关蛋白赤霉素受体GID1L2在GT29中均下调表达, 在Yacheng 71-374中无差异。这表明ABA在甘蔗抗黑穗病过程中可能有重要作用。同Su等[11]鉴定结果一致, 茉莉酸(JA)、水杨酸(SA)、细胞分裂素(CTK)相关蛋白没有被鉴定到, 可能其在早期被诱导, 采样时间点较晚, 表达丰度低。
2.10 共有差异表达蛋白分析
2个甘蔗品种共同差异表达蛋白有83个。其中GT29上调表达蛋白55个, 下调表达蛋白28个; Yacheng 71-374上调表达蛋白48个, 下调表达蛋白35个。对83个共有差异表达蛋白进行COG功能分类, 整体功能类(general function prediction only)蛋白数量最多, 其次是翻译/核糖体结构域生物起源(translation, ribosomal structure and biogenesis)(图3)。表4列出了在GT29中上调表达而在Yacheng 71-374中下调表达的共有差异表达蛋白。
3 讨论
通过iTRAQ技术鉴定到感黑穗病后甘蔗品种GT29和Yacheng 71-374中有定量信息蛋白分别为1429个和1576个, 差异表达蛋白分别为290个和125 个, 远多于本课题组此前利用双向电泳技术鉴定到甘蔗品种ROC22感黑穗病后差异表达蛋白数(18个)[9], 证明了iTRAQ技术的高通量。其中抗病品种GT29的差异表达蛋白多于感病品种Yacheng 71-374, GT29在KEGG富集到的代谢通路也更多, 可能感病后抗病品种的免疫调节机制更为复杂, 涉及的调控通路网更广。对鉴定到的所有蛋白进行GO注释, 在生物过程、分子功能、细胞组分中不同功能蛋白数分布情况与苏亚春等[7]结果一致, 但对差异表达蛋白富集分析结果则有所不同, 可能不同时期不同品种甘蔗对黑穗病应答机制有所差异。
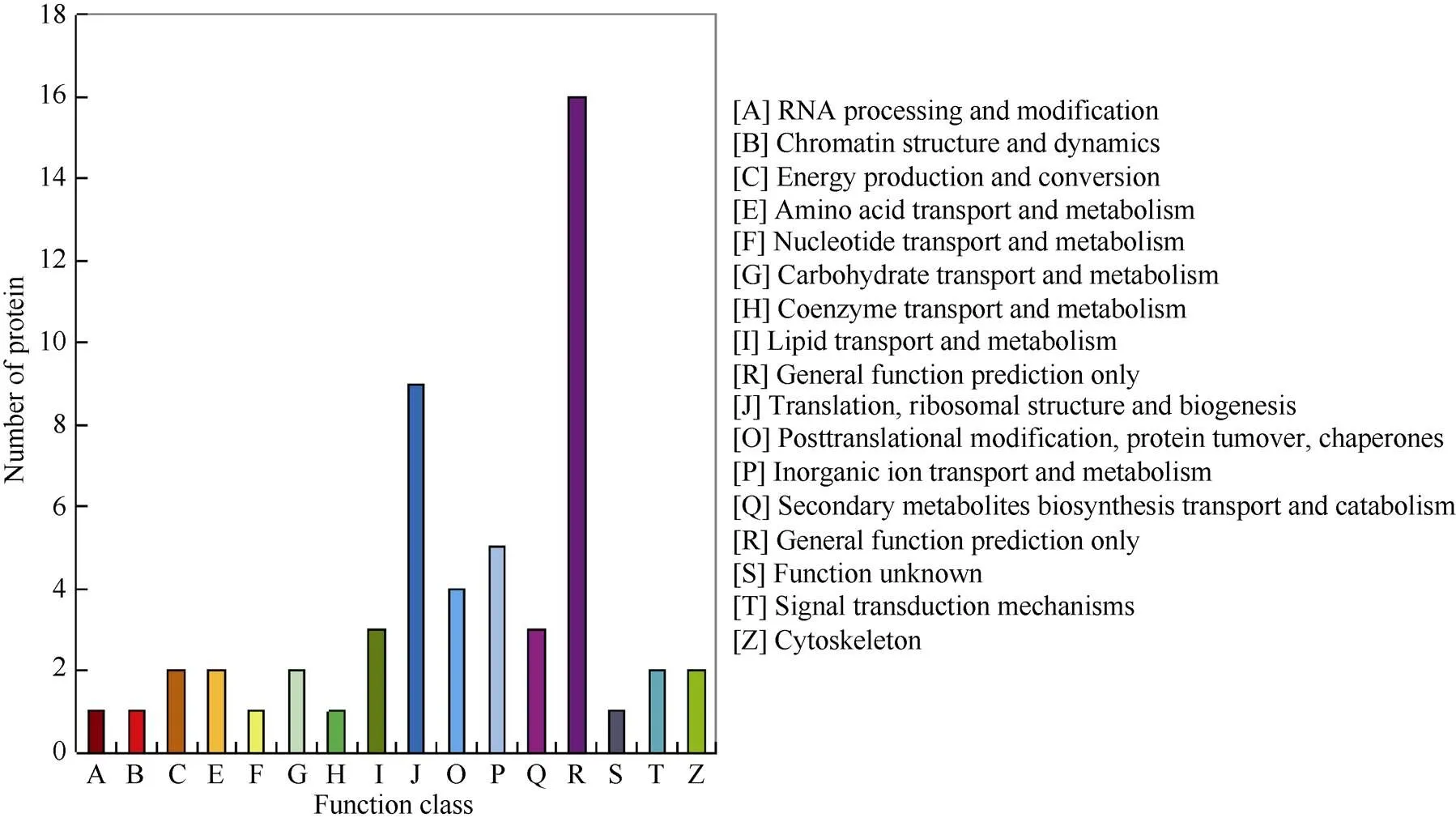
图3 共有差异表达蛋白COG功能分类

表4 在GT29中上调表达、在Yacheng 71-374中下调表达的共有差异表达蛋白
为上调表达,¯为下调表达。is up-regulated and¯is down-regulated.
绿色植物通过光合作用合成有机物质和获取能量, 是植物重要的生理活动。许多研究表明, 植物在受到生物或非生物胁迫后, 通常光合能力受到影响, 相关参数与对照比发生变化[32-36]。沈喜等[37]通过研究感病品种和抗病品种小麦感染条锈病菌后0~12 d叶绿素含量变化显示, 感病品种的叶绿素含量一直在下降, 而抗病品种叶绿素含量呈先下降后上升趋势, 推测抗病品种通过叶绿素机制来抵御病原菌入侵。本研究发现, 与光合作用相关差异表达蛋白在2个甘蔗品种中大多为上调表达, 且抗性品种上调表达蛋白数多于感病品种。差异表达蛋白参与了光合作用的各个方面, 如光捕获、光传递、电子传递、维持蛋白复合体结构稳定、能量转换等。推测光合作用在甘蔗感黑穗病后期免疫机制中可能起重要作用, 上调表达蛋白可能有利于修复受损的光合系统, 进而维护植物生长, 提高对病原菌抗性。
植物对各种胁迫的反应之一是ROS的快速生成,而过量的ROS会氧化蛋白质、脂质和核酸, 从而导致细胞结构的损伤[38]。对于维持细胞体内的ROS平衡, 控制过多ROS带来的毒性影响, 植物建立了多重的抗氧化酶系统[39]。本研究抗性品种GT29中鉴定到POD差异表达蛋白有11个上调表达, 4个下调表达, 感病品种Yacheng 71-374有10个上调表达, 3个下调表达。Su等[11]对甘蔗感病初期研究结果显示, ROS相关蛋白并没有表现出与抗性显著相关, 其中POD在抗性品种Yacheng 05-179中有6个上调表达蛋白, 6个下调表达蛋白, 感病品种ROC22中10个蛋白全部为上调表达, 尽管如此, 本研究与其相似的是POD差异表达蛋白均在抗氧化酶系统中占多数, 且上调表达蛋白在4个品种中数量均较多。暗示POD在甘蔗应答黑穗病过程中可能有重要作用。POD和CAT能将H2O2分解成无毒害的H2O和O2, 两者在一定强度的环境胁迫下有互补的作用, 后期CAT活性减弱, POD活性增强[40]。
植物激素将生长和环境信号整合到一个精细的信号网络中, 在植物的生长发育中起着至关重要的作用, 其不但能调控植物株型的形成, 而且能使植物适当地应对潜在的压力[41]。对激素相关差异蛋白分析, 发现主要为ABA相关蛋白[蛋白磷酸酶2C(PP2C)、9-顺式-环氧类胡萝卜素双加氧酶(NCED)]。NCED是高等植物ABA生物合成途径的关键酶[42], 当植物被诱导产生ABA后, PYR/PYL/ RCAR作为ABA信号受体, 与PP2C家族的A类蛋D结合, 释放SnRK2s类激酶, 激活ABA信号反应[43]。ABA相关差异蛋白均为上调表达, GT29蛋白个数多于Yacheng 71-374, 显示ABA对甘蔗抗黑穗病有重要作用, 这与Que等[44]转录组分析结果一致。ABA能促进植物的防御, 并参与复杂的协同和拮抗相互作用网络, 它在抗病中的作用取决于病原菌的类型、进入宿主的具体方式以及防御反应的时间和植物组织的类型[45]。ABA在植物抗病中的作用已有大量报道[46-49]。本实验发现钙信号通路在一定程度上参与了甘蔗抗病过程, Ca2+信号作为第二信使在ABA信号转导过程中也发挥着重要作用[50], 可能参与了ABA的调控。
2个甘蔗品种共有差异表达蛋白经COG功能分类显示在整体功能类蛋白数最多, 其中在GT29上调表达而在Yacheng 71-374下调表达的蛋白除与光合作用和活性氧代谢相关外, 还参与了多种途径与功能。核糖体蛋白(c62874_g2; orf1、c60900_g1; orf1、c61922_g1; orf1、c53669_g1; orf1)除参与蛋白质的生物合成外, 还具有转录、RNA加工、DNA修复等功能[51]。NAD(P)H-醌氧化还原酶催化醌到氢醌的有益双电子还原, 防止在分子氧存在下通过半醌类的氧化还原反应形成活性氧物质[52]。过氧化氢酶是一种有效的活性氧清除剂, 主要功能是消除在发育过程中生物/非生物胁迫产生的过量的过氧化氢, 同样参与ROS代谢。蛋白c54219_g1; orf2、c70387_g2; orf1、c58104_g1; orf1、c54911_g1; orf1为光合作用相关蛋白。C4磷酸烯醇式丙酮酸羧化酶(c42694_g1; orf1、c42694_g2; orf1、c53868_g3; orf1)在C4植物中作为C4二羧酸循环的关键酶, 起着固定大气中CO2并作为CO2泵而提高光合效率的作用; 同时可以调节细胞pH, 保持离子平衡[53]。2A型丝氨酸/苏氨酸蛋白磷酸酶在植物代谢和信号转导中发挥着多种调控作用[54]。γ-生育酚甲基转移酶是天然维生素E合成途径中的关键酶之一, 其表达及活性对决定植物维生素E的组成起着重要作用[55]。已有研究表明光合器官的维生素E在逆境如强光、干旱、重金属、高温、低温条件下大幅升高[56]。
苯丙烷类代谢相关蛋白及一些酶(GPX、APX、SOD、GST、CAT)活性、激素(IAA、ETH、GA)在前人研究中与植物抗病相关[57], 表现为抗病品种在感病后表达量均比感病对照显著增加。但在本实验中没有发现参与甘蔗的抗病过程, 可能与采样时间有关。前人研究多为感病初期, 而各个代谢途径产生作用的时间与环境、植物、病原菌自身均有联系, 即使在十几个小时内也是不断变化的[58], 这是一个复杂的代谢过程。
4 结论
感黑穗病后抗病甘蔗品种GT29的差异表达蛋白数多于感病品种Yacheng 71-374, GT29在KEGG富集到的代谢通路也更多。光合作用通路、ROS途径、ABA、钙信号通路相关蛋白在2个品种中多为上调表达, 且GT29的上调表达蛋白数多于Yacheng 71-374, 可能参与甘蔗后期对黑穗病的应答。植物抗病是一个复杂的过程, 需要多种功能与途径参与调控。
[1] Martinez M, Medina I, Naranjo S, Rodriguez C, Armas R, Pinon D, Vicente C, Legaz M E. Changes of some chemical parameters, involved in sucrose recovery from sugarcane juices, related to the susceptibility or resistance of sugarcane plants to smut ()., 2000, 102: 445–448.
[2] Su Y C, Wang Z Q, Xu L P, Peng Q, Liu F, Li Z, Que Y X. Early selection for smut resistance in sugarcane using pathogen proliferation and changes in physiological and biochemical indices., 2016, 7: e84426.
[3] Lloyd H L. Chemical assay potentially suitable for determination of smut resistance of sugarcane cultivars., 1983, 67: 1103–1105.
[4] Singh A P, Lal R, Solomon S. Changes in ascorbic acid content in sugarcane affected with smut fungus (Syd.)., 2002, 4: 72–73.
[5] Santiago R, Quintana J, Rodríguez S, Díaz E M, Legaz M E, Vicente C. An elicitor isolated from smut teliospores () enhances lignin deposition on the cell wall of both sclerenchyma and xylem in sugarcane leaves., 2010, 42: 2867–2881.
[6] 莫凤连, 杨丽涛, 潘如科, 宋修鹏, 李杨瑞. 甘蔗黑穗病菌胁迫对甘蔗内源激素含量的影响. 南方农业学报, 2012, 43: 1676–1681. Mo F L, Yang L T, Pan R K, Song X P, Li Y R. Changes of endogenous hormone content in sugarcane under smut pathogen stress., 2012, 43: 1676–1681 (in Chinese with English abstract).
[7] 苏亚春. 甘蔗应答黑穗病菌侵染的转录组与蛋白组研究及抗性相关基因挖掘. 福建农林大学博士学位论文, 福建福州, 2014. Su Y C. Transcriptomics and Proteomics of Sugarcane Response toInfection and Mining of Resistance-Related Genes. PhD Dissertation of Fujian Agriculture and Forestry University, Fuzhou, Fujian, China, 2014 (in Chinese with English abstract).
[8] Leila P, Peters G, Carvalho M B, Vilhena S, Creste R A, Azevedo C B, Monteiro V. Functional analysis of oxidative burst in sugarcane smut-resistant and smut-susceptible genotypes., 2017, 245: 749–764.
[9] 宋修鹏. 黑穗病菌侵染初期甘蔗幼苗光合生理变化及基因差异表达. 广西大学博士学位论文, 广西南宁, 2014. Song X P. Effects of Smut Pathogen on Photosynthetic Characteristics and Gene Differential Expression in Sugarcane at Seedling Stage.PhD Dissertation of Guangxi University, Nanning, Guangxi, China, 2014 (in Chinese with English abstract).
[10] Barnabas L, Ashwin N M R, Kaverinathan K. Proteomic analysis of a compatible interaction between sugarcane and., 2016, 16: 1111–1122.
[11] Su Y C, Xu L P, Wang Z, Peng Q, Yang Y T, Chen Y, Que Y X. Comparative proteomics reveals that central metabolism changes are associated with resistance againstin sugarcane., 2016, 17: 800.
[12] Que Y X, Xu L P, Lin J W, Ruan M H, Zhang M Q, Chen R K. Differential protein expression in sugarcane during sugarcane-interaction revealed by 2-de and maldi-tof-tof/ms., 2011: 989016.
[13] 钟云.诱导的柑橘转录组学及蛋白组学研究. 湖南农业大学博士学位论文, 湖南长沙, 2012 Zhong Y. Proteomics of Citrus Induced by. PhD Dissertation of Hunan Agricultural University, Changsha, Hunan, China, 2012 (in Chinese with English abstract)
[14] 唐成, 陈露, 安敏敏, 孟丹, 杨立明, 罗玉明. 水稻幼苗叶片应答稻瘟病侵染的差异蛋白谱分析. 淮阴师范学院学报, 2014, 13: 322–328. Tang C, Chen L, An M M, Meng D, Yang L M, Luo Y M. Proteomic analysis reveals an intimate protein pathways provoked by blast in rice seedling leaves., 2014, 13: 322–328 (in Chinese with English abstract).
[15] Marsh E, Alvarez S, Hicks L M, Barbazuk B W, Qiu W P, Kovacs L, Schachtman D. Changes in protein abundance during powdery mildew infection of leaf tissues of Cabernet Sauvignon grapevine (L.)., 2010, 10: 2057–2064.
[16] 张荣华, 何红, 张革民, 刘海斌, 李杨瑞, 方锋学, 宋焕忠, 方位宽, 闭少玲. 宿根性特强甘蔗新品种桂糖29号的选育. 中国糖料, 2011, (1): 1–4. Zhang R H, He H, Zhang G M, Liu H B, Li Y R, Fang F X, Song H Z, Fang W K, Bi S L. Breeding of new sugarcane variety Guitang 29 with high ratoon ability., 2011, (1): 1–4 (in Chinese with English abstract).
[17] 高轶静, 张革民, 张荣华, 宋焕忠, 罗霆, 段维兴, 贤武, 廖江雄, 周会, 游建华. 甘蔗优良新品种(系)的黑穗病抗性鉴定. 中国糖料, 2013, (2): 25–26. Gao Y J, Zhang G M, Zhang R H, Song H Z, Luo T, Duan W X, Xian W, Liao J X, Zhou H, You J H. Evaluation of resistance to smut disease in new sugarcane varieties and breeding lines.2013, (2): 25–26 (in Chinese with English abstract).
[18] 熊国如, 张树珍. 甘蔗黑穗病的研究. 农业灾害研究, 2012, 2(4): 8–10. Xiong G R, Zhang S Z. Study on sugarcane smut., 2012, 2(4): 8–10 (in Chinese with English abstract).
[19] 林丽, 张新成, 李杨瑞, 梁俊. 甘蔗器官固氮酶活性及其对接种固氮菌的响应. 西北植物学报, 2008, 28: 2472–2477. Lin L, Zhang X C, Li Y R, Liang J. Changes of nitrogenase activity in sugarcane (L.) and its response to innoculate nitrogen fixation bacteria., 2008, 28: 2472–2477 (in Chinese with English abstract).
[20] 彭浩, 林文芳, 朱学艺. 叶绿体蛋白质组研究进展. 西北植物学报, 2008, 28: 194–203. Peng H, Lin W F, Zhu X Y. Research progress in chloroplast proteome., 2008, 28: 194–203 (in Chinese).
[21] 邱念伟, 王颖. 光合作用光反应过程中的物质与能量转换. 生物学教学, 2011, 36(2): 71–74 Qiu N W, Wang Y. Substance and energy conversion during photosynthesis photoreaction., 2011, 36(2): 71–74 (in Chinese).
[22] 苟萍, 索菲娅, 马东建. 高等植物铁氧还蛋白的结构与功能. 生命的化学, 2007, 27: 51–53. Gou P, Suo F Y, Ma D J. Structure and function of ferredoxin in higher plants., 2007, 27: 51–53 (in Chinese).
[23] 翟玉山, 邓宇晴, 董萌, 徐倩, 程光远, 彭磊, 林彦铨, 徐景升. 甘蔗捕光叶绿素结合蛋白基因的克隆及表达. 作物学报, 2016, 42: 1332–1341. Zhai Y S, Deng Y Q, Dong M, Xu Q, Cheng G Y, Peng L, Lin Y Q, Xu J S. Cloning and characterization of light harvesting chlorophyll-binding protein coding gene () in sugarcane., 2016, 42: 1332–1341 (in Chinese with English abstract).
[24] Mayfield S P, Bennoun P, Rochaix J D. Expression of the nuclear encoded OEE1 protein is required for oxygen evolution and stability of photosystem II particles in Chlamydomonas reinhardtii., 1987, 6: 313–318.
[25] Medentsev A G, Arinbasarova A I, Aimenko V K. Adaptation of the phytopathogenic fungusdecemcellulare to oxidative stress., 2001, 70: 34–38.
[26] 余晓丛, 娜仁, 张少英. 钙信号在植物抗病性中的作用研究进展. 中国农学通报, 2012, 28(3): 12–16. Yu X C, Na R, Zhang S Y. Research progress about calcium signal involved in plant resistance to disease., 2012, 28(3): 12–16 (in Chinese with English abstract).
[27] 李丽花, 张瑞杰, 姚远丽, 朱璇. 24-表油菜素内酯调控苯丙烷代谢增强杏果实抗病性的研究. 现代食品科技, 2017, 33(9): 71–76. Li L H, Zhang R J, Yao Y L, Zhu X. 24-epibrassinolide enhances disease resistance in apricot fruits via regulation of phenylpropanoid metabolism.2017, 33(9): 71–76 (in Chinese with English abstract).
[28] 杜翔宇, 刘春燕, 吴琼, 蒋洪蔚, 辛大伟, 陈庆山, 胡国华. 大豆苯丙氨酸代谢途径关键酶基因的挖掘定位及结构分析. 大豆科学, 2012, 31: 178–183. Du X Y, Liu C Y, Wu Q, Jiang H W, Xin D W, Chen Q S, Hu G H. Gene mining and structure analysis of key enzyme genes on phenylalanine acid metabolic pathway in soybean., 2012, 31: 178–183 (in Chinese with English abstract)
[29] Liu D X, Fan C S, Tao J H. Integration ofaroG-pheA tandem genes intotyrA locus and its effect on L-phenylalanine biosynthesis., 2004, 10: 3683–3687.
[30] 张烨. 柠条锦鸡儿咖啡酰辅酶A-O-甲基转移酶基因cDNA和gDNA全长克隆及生物信息学分析. 内蒙古农业大学硕士学位论文, 内蒙古呼和浩特, 2011. Zhang Y. Full-length cDNA and Genomic DNA Cloning of Caffeoyl Coenzyme A-O-methyltransferase from Caragana Korshinkii Kom and Its Bioinformatics Analysis.MS Thesis of Inner Mongolia Agricultural University, Hohhot, Inner Mongolia, China, 2011 (in Chinese with English abstract).
[31] 王志卫, 贝学军, 朱世平. 植物激素在植物抗病过程中的作用研究进展. 安徽农业科学, 2011, 39: 9035–9038. Wang Z W, Bei X J, Zhu S P. Recent advances in phytohormone regulated plant resistance to pathogens.,2011, 39: 9035–9038 (in Chinese with English abstract).
[32] 赵琴, 潘静, 曹兵, 宋丽华.气温升高与干旱胁迫对宁夏枸杞光合作用的影响. 生态学报, 2015, 35: 6016–6022. Zhao Q, Pan J, Cao B, Song L H. Effects of elevated temperature and drought stress on photosynthesis of., 2015, 35: 6016–6022 (in Chinese with English abstract)
[33] 庞杰, 张凤兰, 郝丽珍, 杨忠仁, 赵鹏.沙芥幼苗叶片解剖结构和光合作用对干旱胁迫的响应. 生态环境学报, 2013, 22: 575–581. Pang J, Zhang F L, Hao L Z, Yang Z R, Zhao P. Effect of drought stress on anatomical structure and photosynthesis of(L.)Gaertn. leaves in seedling., 2013, 22: 575–581 (in Chinese with English abstract).
[34] 孙璐, 周宇飞, 李丰先, 肖木辑, 陶冶, 许文娟, 黄瑞冬. 盐胁迫对高粱幼苗光合作用和荧光特性的影响. 中国农业科学, 2012, 45: 3265–3272. Sun L, Zhou Y F, Li F X, Xiao M J, Tao Y, Xu W J, Huang R D. Impacts of salt stress on characteristics of photosynthesis and chlorophyll fluorescence of sorghum seedlings., 2012, 45: 3265–3272 (in Chinese with English abstract).
[35] 马富举, 李丹丹, 蔡剑, 姜东, 曹卫星, 戴廷波.干旱胁迫对小麦幼苗根系生长和叶片光合作用的影响. 应用生态学报, 2012, 23: 724–730. Ma F J, Li D D, Cai J, Jiang D, Cao W X, Dai T B. Responses of wheat seedlings root growth and leaf photosynthesis to drought stress., 2012, 23: 724–730 (in Chinese with English abstract).
[36] 徐田军, 董志强, 兰宏亮, 裴志超, 高娇, 解振兴.低温胁迫下聚糠萘合剂对玉米幼苗光合作用和抗氧化酶活性的影响. 作物学报, 2012, 38: 352–359. Xu T J, Dong Z Q, Lan H L, Pei Z C, Gao J, Xie Z X. Effects of PASP-KT-NAA on photosynthesis and antioxidant enzyme activities of maize seedlings under low temperature stress., 2012, 38: 352–359 (in Chinese with English abstract).
[37] 沈喜, 李红玉, 贾秋珍, 冯汉青, 李敏权, 梁厚果.条锈病对小麦(L.)叶片光合功能及光合功能蛋白D1表达的影响. 生态学报, 2008, 8: 669–676. Shen X, Li H Y, Jia Q Z, Feng H Q, Li M Q, Liang H G. Influence of wheat (L.) stripe rust infection on photosynthetic function and expression protein D1 of what leaves., 2008, 8: 669–676 (in Chinese with English abstract).
[38] Medentsev A G, Aiu A, Aimenko V K. Adaptation of the phytopathogenic fungusdecemcellulare to oxidative stress., 2001, 70: 26–30.
[39] Mittle R, Vanderauwera S, Gollery M, Breusegem F V. Reactive oxygen gene network of plants., 2004, 9: 490–498.
[40] 张永志, 赵首萍, 徐明飞, 王钢军, 郑纪慈. Pb胁迫对番茄幼苗抗氧化酶系统的影响.浙江农业科学, 2009, 1: 452–456 Zhang Y Z, Zhao S P, Xu M F, Wang G J, Zheng J C. Effect of Pb stress on antioxidant enzyme system in tomato seedlings., 2009, 1: 452–456 (in Chinese with English abstract).
[41] Kazan K, Manners J M. Linking development to defense: auxin in plant–pathogen interactions., 2009, 14: 373–382.
[42] 朱海生, 李永平, 花秀凤, 温庆放. 草莓9-顺式-环氧类胡萝卜素双加氧酶基因的克隆及表达分析. 园艺学报, 2012, 39: 40–48. Zhu H S, Li Y P, Hua X F, Wen Q F. Cloning and expression analysis of 9--epoxycarotenoid dioxygenase genein strawberry., 2012, 39: 40–48 (in Chinese with English abstract).
[43] 胡帅, 王芳展, 刘振宁, 刘亚培, 余小林. PYR/PYL/RCAR蛋白介导植物ABA的信号转导. 遗传, 2012, 34: 560–572 Hu S, Wang F Z, Liu Z N, Liu Y P, Yu X L. ABA signaling mediated by PYR/PYL/RCAR inplants., 2012, 34: 560–572 (in Chinese with English abstract).
[44] Que Y X, Su Y C, Guo J L, Wu Q B, Xu L P. A global view of transcriptome dynamics during,, challenge in sugarcane by RNA-seq., 2014, 9: e106476.
[45] Ton J, Flors V, Mauchmani B. The multifaceted role of ABA in disease resistance., 2009, 14: 310–317.
[46] Fraser R S S. Are ‘pathogenesis-related’ proteins involved in acquired systemic resistance of tobacco plants to tobacco mosaic virus., 1982, 58: 305–313.
[47] Fujita M, Fujita Y, Noutoshi Y. Crosstalk between abiotic and biotic stress responses: a current view from the points of convergence in the stress signaling networks., 2006, 9: 436–442.
[48] Koga H, Dohi K, Mori M. Abscisic acid and low temperatures suppress the whole plant-specific resistance reaction of rice plants to the infection of., 2004, 65: 3–9.
[49] 周金鑫, 胡新文, 张海文, 黄荣峰. ABA在生物胁迫应答中的调控作用. 农业生物技术学报, 2008, 16: 169–174. Zhou J X, Hu X W, Zhang H W, Huang R F. Regulatory role of ABA in plant response to biotic stresses., 2008, 16: 169–174 (in Chinese with English abstract).
[50] 钱万强. 松材线虫侵染后马尾松体内的苯丙烷代谢研究. 中南林业科技大学硕士学位论文, 湖南长沙, 2009. Qian W Q. Phenylpropane Metabolic Research ofInfected byMS Thesis of Central South University of Forestry and Technology, Changsha, Hunan, China, 2009 (in Chinese with English abstract).
[51] Wool I G. Extraribosomal functions of ribosomal proteins., 1996, 21: 164–169.
[52] Vasiliou V, Ross D, Nebert D W. Update of the NAD(P)H: quinone oxidoreductase (NQO) gene family., 2006, 2: 329–335.
[53] 魏绍巍, 黎茵. 植物磷酸烯醇式丙酮酸羧化酶的功能及其在基因工程中的应用. 生物工程学报, 2011, 27: 1702–1710. Wei S W, Li Y. Functions of plant phosphoenolpyruvate carboxylase and its applications for genetic engineering., 2011, 27: 1702–1710 (in Chinese with English abstract).
[54] Yu R M, Zhou Y, Xu Z F. Two genes encoding protein phosphatase 2A catalytic subunits are differentially expressed in rice., 2003, 51: 295–311.
[55] Dellapenna D, Pogson B J. Vitamin synthesis in plants: tocopherols and carotenoid., 2006, 57: 711–738.
[56] 刘宾. 玉米γ-生育酚甲基转移酶基因的分离及其功能分析. 河北大学硕士学位论文, 河北保定, 2007.Liu B. Solating and Functional Analysis of γ-tocopherol Methytransferase Gene in. MS Thesis of Hebei University, Baoding, Hebei, China, 2007 (in Chinese with English abstract).
[57] 关西贞, 张卫东, 田纪春. 小麦近等基因系与白粉病菌互作的生理指标研究. 华北农学报, 2010, 25(1): 217–221. Guan X Z, Zhang W D, Tian J C. Physiological indicators of near-isogenic wheat lines in interaction with powdery mildew.-, 2010, 25(1): 217–221 (in Chinese with English abstract).
[58] 刘守伟, 吴凤芝, 马艳玲. 枯萎病菌对不同抗性黄瓜品种几种酶活性的影响. 植物保护, 2009, 35(1): 82–85.Liu S W, Wu F Z, Ma Y L. Effects of fusarium wilt pathogen on the enzyme activity of cucumber cultivars of different resistance., 2009, 35(1): 82–85 (in Chinese with English abstract).
附表1 GT29差异表达蛋白GO富集分析
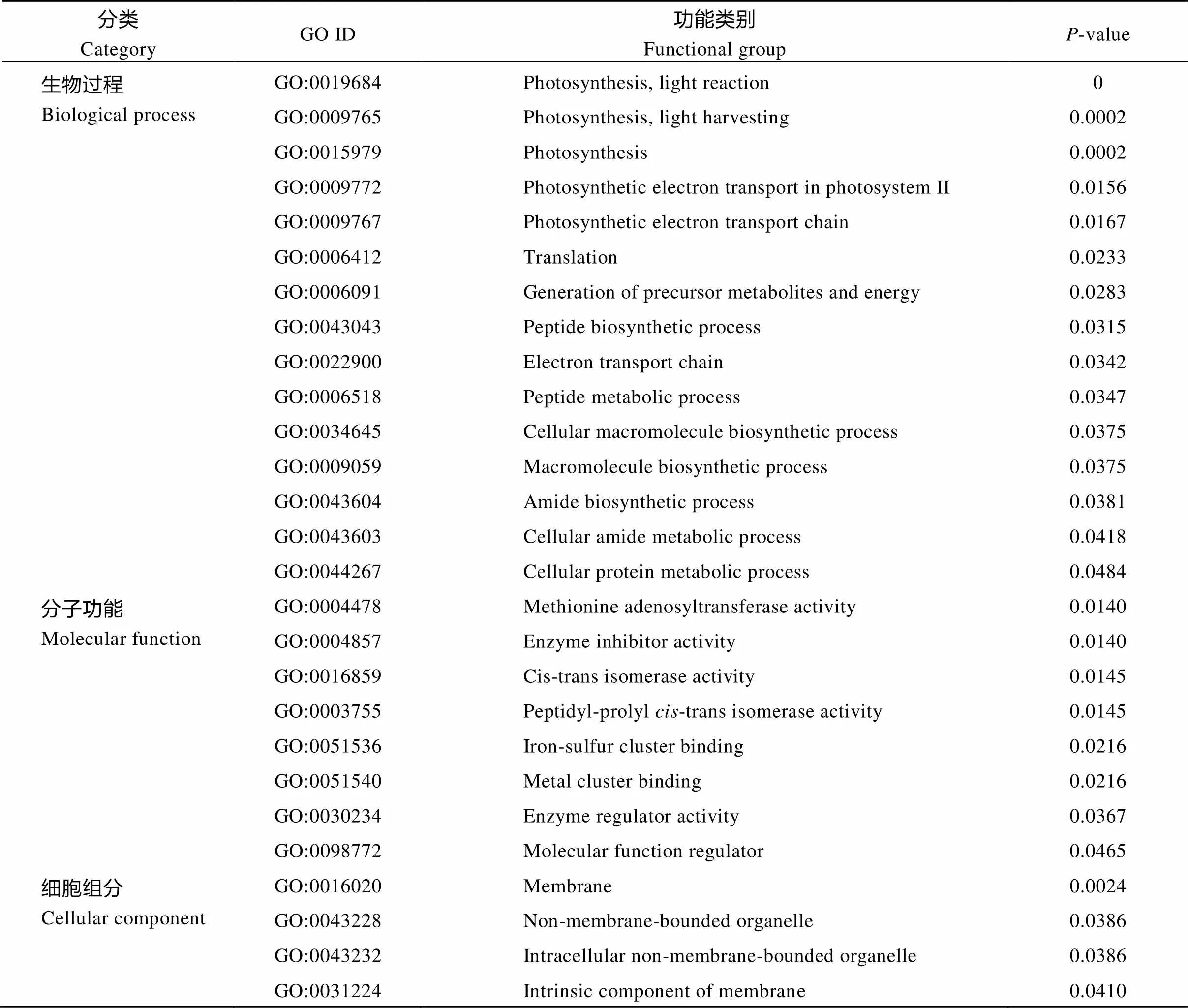
Supplementary table 1 Differentially expressed protein GO enrichment analysis for GT29
附表2 Yacheng 71-374差异表达蛋白GO富集分析
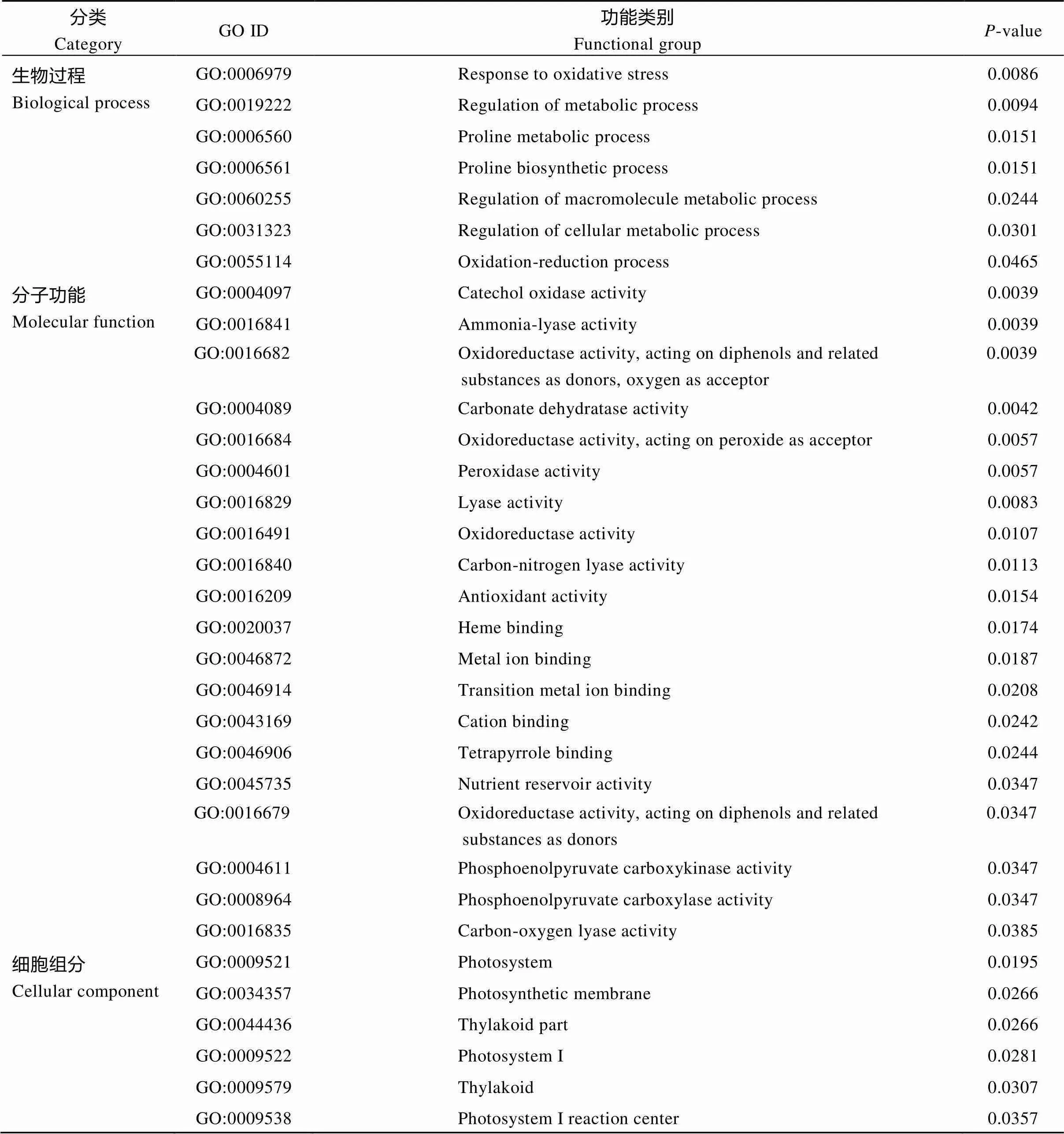
Supplementary table 2 Differentially expressed protein GO enrichment analysis for Yacheng 71-374
Proteomic analysis of sugarcane-interaction based on iTRAQ technique
SONG Qi-Qi1,**, Pratiksha SINGH2,**, Rajesh Kumar SINGH2, SONG Xiu-Peng1, LI Hai-Bi1, NONG You-Ye2, YANG Li-Tao1,2,*, and LI Yang-Rui1,2,*
1Sugarcane Research Center, ChineseAcademy of Agricultural Sciences / Guangxi Academy of Agricultural Sciences / Key Laboratory of Sugarcane Biotechnology and GeneticImprovement (Guangxi), Ministry of Agriculture / Guangxi Key Laboratory of Sugarcane Genetic Improvement, Nanning 530007, Guangxi, China;2AgriculturalCollege, GuangxiUniversity / State Key Laboratory for Conservation and Utilization of Subtropical Agro-bioresources, Nanning 530005, Guangxi, China
Sugarcane smut has become an important disease affecting sugarcane yield and sugar content. In order to investigate the molecular mechanism of sugarcane responding to smut at protein level, the smut-resistant variety GT29 and the smut-susceptible variety Yacheng 71-374 were used in this study. Both varieties were inoculated with the teliospore suspension of smut pathogen by dipping method whereas the control was treated with sterile water. Leaf samples were collected and used for proteomic analysis by iTRAQ technique at 180 days after treatment. The that 1429 proteins presented in GT29 with quantitative information, including 290 differentially expressed proteins with 153 up-regulated and 137 down-regulated; while 1576 proteins in Yacheng 71-374 with quantitative information, including 125 differentially expressed proteins with 55 up-regulated and 70 down-regulated. The number of differentially expressed proteins in resistant variety was higher than that in susceptible variety, and GT29 enriched more metabolic pathways in KEGG, indicating that the immunomodulatory mechanism of the resistant variety may be more complicated, and the regulatory network involved in response was broader. Through the analysis of photosynthesis, antioxidant system, calcium signal, phenylpropane metabolism, hormone related differential expressed protein and co-owned differentially expressed protein, it was found that the photosynthesis pathway, ROS, ABA, calcium signal pathway related protein were up-regulated in both varieties. The up-regulated expressed proteins were more in GT29 than in Yacheng 71-374, which may be involved in resistance response in sugarcane against smut pathogen at later growth stage. Plant disease resistance is a complex process that requires multiple functions and pathways to participate in regulation. The phenylpropanoid metabolic pathway, enzymes GPX, APX, SOD, GST, and CAT, and hormones IAA, ETH, and GA were not found to be involved in the disease resistance process of sugarcane.
sugarcane; smut; iTRAQ; proteomics
2018-01-04;
2018-08-20;
2018-09-26.
10.3724/SP.J.1006.2019.84001
通信作者(Corresponding authors):李杨瑞, E-mail: lyr@gxaas.net; 杨丽涛, E-mail: litao61@hotmail.com
**同等贡献(Contributed equally to this work)
宋奇琦, E-mail: 2632754996@qq.com
本研究由广西八桂学者和特聘专家专项经费项目([2013]3号), 广西科技基地和人才专项(桂科AD17195100), 国家现代农业产业技术体系广西甘蔗创新团队项目(gjnytxgxcxtd-03-01), 广西自然科学基金项目(2015GXNSFBA139060)和广西甘蔗遗传改良重点实验室项目(16-K-02-01)资助。
This study was supported by the Guangxi Special Fund for Scientific Base and Talent (GKAD17195100), the Special Funds for Bagui Scholars and Distinguished Experts in Guangxi (2013-03), Guangxi Sugarcane Innovation Team of National Agricultural Industry Technology System (gjnytxgxcxtd-03-01), Guangxi Natural Science Fundation (2015GXNSFBA139060), and Guangxi Key Laboratory of Sugarcane Genetic Improvement (16-K-02-01).
URL:http://kns.cnki.net/kcms/detail/11.1809.S.20180921.1625.006.html
猜你喜欢
河北科技师范学院学报(2022年2期)2022-08-26
中国糖料(2022年2期)2022-04-06
重庆科技学院学报(自然科学版)(2021年5期)2021-11-10
现代养生·上半月(2021年10期)2021-09-24
中国药剂学杂志(网络版)(2019年4期)2019-07-17
水产科学(2019年3期)2019-05-30
分析化学(2018年4期)2018-11-02
分析化学(2018年7期)2018-09-17
农家科技中旬版(2017年12期)2018-02-26
农家顾问(2016年7期)2016-05-14
