
miR-648靶向Syk对胃癌细胞增殖、凋亡、迁移、侵袭的影响及机制研究
2019-12-13 07:04:20何淑兵左东明张文山刘博智英辉张国栓
现代消化及介入诊疗 2019年11期
何淑兵,左东明,张文山,刘博,智英辉,张国栓
胃癌是起源于胃黏膜上皮的恶性肿瘤,在我国各种恶性肿瘤中发病率居第二位[1]。全球每年新发胃癌病例100多万,死亡约80多万,其发病有明显的地域差别,其中东南亚地区发病率最高,中国每年新发胃癌病例占全球的42%,死亡占35%[2]。因此,深入研究胃癌发生发展的机制,寻找诊断新的标志物和治疗新靶点,对防治胃癌提高人们的生活质量具有重要价值。
近年来研究发现,微小型RNA(microRNA,miRNA)是一种新的基因调控元件,其长度约为22个碱基的单链RNA,不具有编码蛋白的功能,可以使其靶基因降解或阻碍其靶基因的翻译来调控其靶基因的表达[3],从而参与调控细胞增殖、凋亡、干细胞分化、肿瘤血管生成、神经元发育、肿瘤细胞浸润和转移等一系列生命活动。研究发现,通过对胃癌细胞及正常胃黏膜上皮细胞体外培养并提取RNA进行miRNA芯片分析发现,与正常胃黏膜细胞相比miR-648在胃癌细胞中上调表达[4],对淋巴结转移胃癌组织及无淋巴结转移胃癌组织应用qRT-PCR检测RNA芯片测定的miRNA结果,证实miR-648在淋巴结转移胃癌组织中明显上调[5],但miR-648对胃癌细胞生物行为的影响及其具体分子机制目前还未见报道。因此,本课题就miR-648对胃癌细胞的生物学影响及其具体分子机制进行研究,以期为胃癌的早期诊断和治疗提供新思路。
1 材料与方法
1.1 材料
胃癌SGC-7901、MGC-803细胞系和正常胃黏膜上皮GES-1细胞购于中国科学院典型培养物宝藏委员会细胞库;胎牛血清(fetal bovine serum,FBS)和DMEM试剂购自Gibco;Trizol总RNA提取试剂购于Invitrogen;RT Profiler PCR Array Kit购于Qiagen;Fast Start Universal SYBR Green Master购于Roche;胰蛋白酶购于Sigma;抗β-actin 抗体购于杭州联科生物科技有限公司;Transwell小室购于Millipore;CCK-8购于Dojindo;细胞凋亡试剂盒购于南京凯基公司。
1.2 方法
1.2.1 细胞培养 采用含10% FBS的DMEM培养基体外培养细胞,置于37 ℃、5% CO2培养箱里培养。选择对数生长期细胞进行实验。
1.2.2 real-time PCR法检测RNA的表达 根据Trizol法提取总RNA,按照PrimeScriptTMRT Master Mix试剂盒说明书进行操作合成cDNA。MiR-648-F: 5′-GGAAGTGTGCAGGGC-3′, MiR-648-R-5′-AGTGCGTGTCGTGGAGTC-3′,脾酪氨酸激酶Syk-F:5′-TTTCGGACTTTCCAAAGCACTGCG-3′, Syk-R:5′-ACTCCAAAGCTCCAGACATCCTGT-3′。U6作为内参,U6-F: 5 ′-GCTTCGGCAGCACATATACTAAAAT-3 ′;U6-R:5′-CGCTT-CACGAATTTGCGTGTCAT-3′。根据SYBR Green I PCR 检测试剂盒说明书在ABI7500荧光定量PCR仪中进行PCR扩增反应,反应条件:95 ℃预变性30 s,95 ℃变性5 s,60 ℃退火32 s,70 ℃延伸30 s,循环40次。溶解曲线分析:95 ℃ 15 s,60 ℃ 30 s,95 ℃ 15 s。2-ΔΔCt法测定各基因mRNA的相对表达水平。每个样品孔重复检测3次,取均值。
1.2.3 Western blot 检测相关蛋白的表达 收集各组细胞,用RIPA细胞裂解液裂解提取各组总蛋白并用BCA法定量。取适量蛋白上样后进行SDS-PAGE,电转至PVDF膜后加入5%脱脂奶粉封闭液室温封闭2 h,孵育相应一抗,4 ℃条件下孵育过夜,洗膜室温孵育相应二抗2 h,洗膜后用化学发光试剂盒对PVDF进行曝光显影,以(-actin为内参,后采用成像扫描分析系统测定条带的灰度值。
1.2.4 细胞转染 收集对数生长期的胃癌SGC-7901细胞,适当密度接种到6孔上,待细胞融合到70%时,按照LipofectmineTM2000转染试剂盒说明书将干扰miR-648的模拟物anti-miR-648和其阴性对照anti-miR-NC转染至SGC-7901细胞中,分别标记为anti-miR-648组和anti-miR-NC组。转染4 h后换为含血清的新鲜培养基继续培养,并用RT-PCR检测转染效果。后续实验中将SGC-7901随机分为miR-648组(转染过表达miR-648模拟物)、miR-NC(转染过表达miR阴性对照)、anti-miR-648+si-Syk组(干扰miR-648表达的模拟物和沉默Syk的siRNA共转染)、anti-miR-648+si-NC组(干扰miR-648表达的模拟物和沉默Syk的阴性对照共转染)四组,采用上述方法转染后进行后续实验。
1.2.5 CCK-8法检测细胞活力 取对数生长期的细胞悬液,按每孔2×103个接种到96孔板,待细胞贴壁后每孔加入CCK-8溶液15 μL,置于37 ℃、5% CO2培养箱重培养2 h,用酶标仪检测490 nm的吸光度。
1.2.6 流式细胞术检测细胞凋亡 收集各组待测细胞,离心漂洗去上清,根据Annexin V-FITC 试剂盒处理细胞,简述步骤如下:缓冲液重悬细胞摇匀,避光孵育15 min后加入Annexin V-FITC, 再次避光孵育15 min后采用流式细胞仪检测细胞的凋亡情况。
1.2.7 Transwell法检测细胞迁移和侵袭 细胞迁移实验如下:Transwell小室在铺板前30 min浸润在无血清培养基中,细胞消化后计数,每孔2×103个,小室内无血清,培养板内有血清,培养24 h后5%多聚甲醛固定15 min,PBS冲洗后结晶紫染色10 min,PBS冲洗后用倒置显微镜拍照。细胞侵袭实验在实验前用20 mg/L Matrigel 1∶3稀释包被Transwell小室底部膜上室面,4 ℃风干后步骤同细胞迁移实验。
1.3 统计学处理

2 结果
2.1 胃癌细胞中miR-648表达上调、Syk表达下调
图1和表1显示,与正常胃黏膜上皮细胞GES-1细胞相比,胃癌SGC-7901、MGC-803细胞中miR-648的表达量明显上调,Syk mRNA和蛋白的表达量明显下调(P<0.05)。
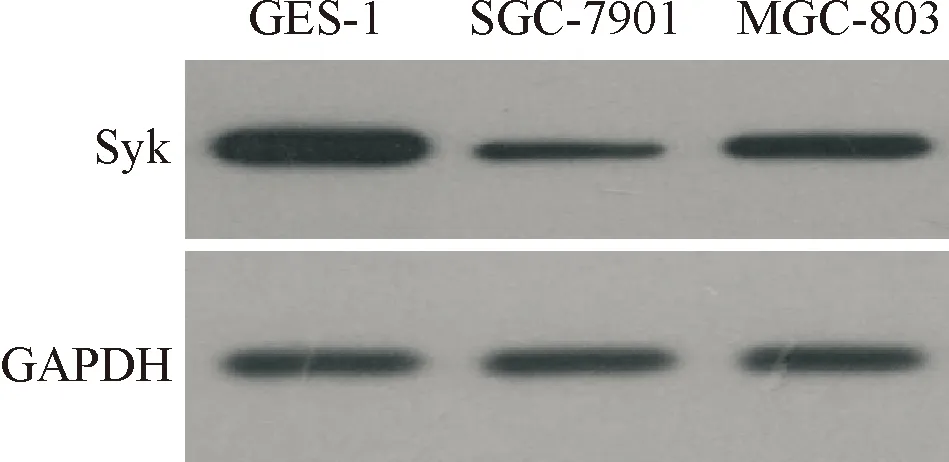
图1Syk蛋白的表达
2.2 抑制miR-648表达抑制了胃癌SGC-7901细胞增殖并促进其细胞凋亡
图2显示,与anti-miR-NC组相比,anti-miR-648组细胞的凋亡率和Bax蛋白的表达量显著上升,Bcl-2蛋白的表达量显著下降(P<0.05)。表2显示,与anti-miR-NC组相比,anti-miR-648组胃癌SGC-7901细胞中miR-648的表达量显著下降(P<0.05),可见,成功转染了抑制miR-648表达的胃癌SGC-7901细胞。转染24 h后,anti-miR-648组与anti-miR-NC组的细胞活性无显著差异,转染48 h和72 h后, anti-miR-648组细胞活性较anti-miR-NC组显著下降;anti-miR-648组较anti-miR-NC组Cyclin D1的表达量明显下降,P21蛋白的表达量明显上升(P<0.05)。

表1miR-648和Syk在 SGC-7901、MGC-803细胞和GES-1细胞中的表达
注:与GES-1组比较,*P<0.05
2.3 抑制miR-648表达抑制了胃癌SGC-7901细胞迁移和侵袭
图3和表3显示,与anti-miR-NC组相比,anti-miR-648组细胞迁移和侵袭的数目显著下降,E-cadherin蛋白的表达量显著升高,MMP-2蛋白的表达量显著下降(P<0.05)。可见,抑制miR-648的表达对胃癌SGC-7901细胞的迁移和侵袭具有抑制作用。
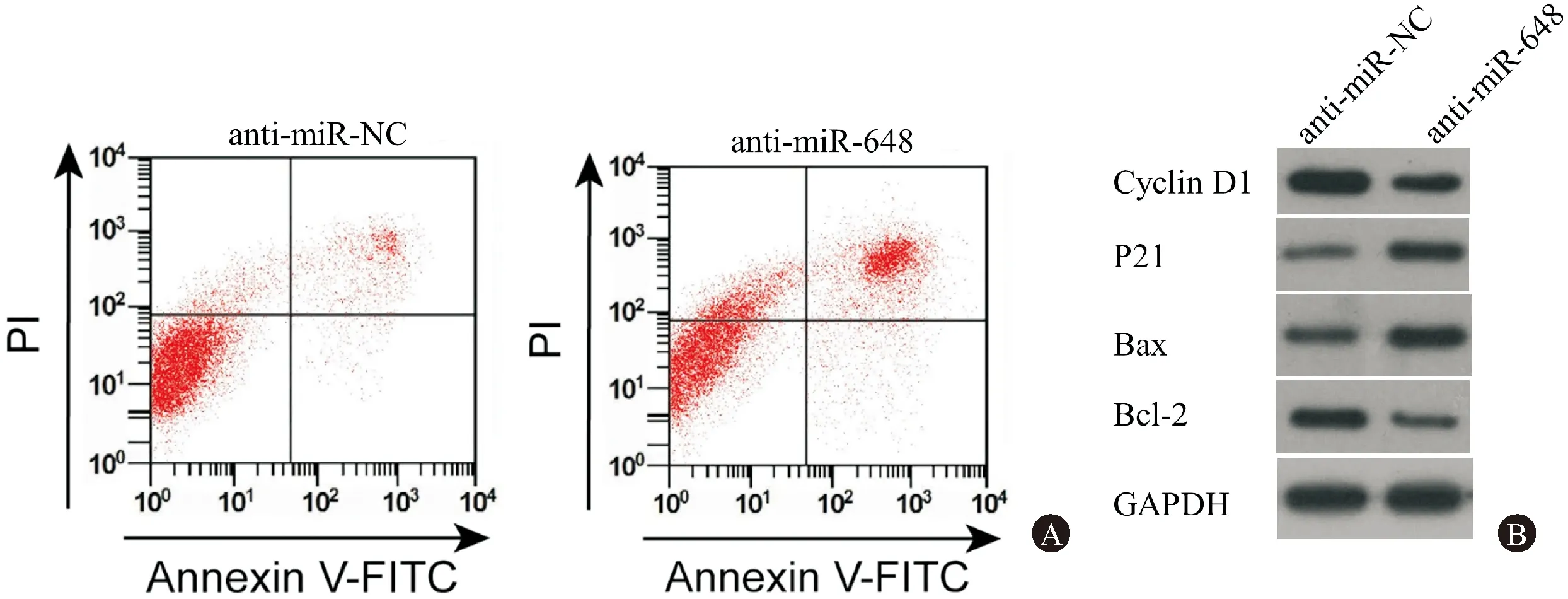
图2抑制miR-648对胃癌细胞SGC-7901凋亡及增殖、凋亡蛋白表达的影响

表2抑制miR-648对胃癌细胞SGC-7901增殖、凋亡的影响

图3抑制miR-648对胃癌细胞SGC-7901迁移、侵袭及相关蛋白表达的影响
2.4 miR-648可靶向抑制Syk的表达
图4A为TargetScan(http://www.targetscan.org/)软件预测结果示意图,表明miR-648与Syk存在互补结合位点。表 4为双荧光素酶报告基因检测结果,同miR-NC与WT-Syk共转染相比,miR-648与WT-Syk共转染后,SGC-7901细胞的荧光素酶活性显著下降(P<0.05);但miR-NC、miR-648分别与MUT-Syk共转染后,SGC-7901细胞的荧光素酶活性差异无统计意义(P<0.05),可见miR-648对Syk具有靶向作用。表5显示,与miR-NC组相比,miR-648组中Syk mRNA和蛋白的表达量显著下降(P<0.05);相反,anti-miR-648组中Syk mRNA和蛋白的表达量较anti-miR-NC组显著升高(P<0.05),可见miR-648可负向调控Syk的表达。
表3抑制miR-648对胃癌细胞SGC-7901迁移、侵袭的影响

图4Syk的3′UTR中含有miR-648的互补核苷酸链
2.5 沉默Syk可逆转抑制miR-648对胃癌细胞SGC-7901增殖、凋亡、迁移和侵袭的影响
图5和表6-7显示,与 anti-miR-NC组相比,anti-miR-648组中Cyclin D1、Bcl-2、MMP-2蛋白的表达量明显下降,Syk、P21、Bax、E-cadherin蛋白的表达量明显上升;转染24 h后,anti-miR-648组与anti-miR-NC组的细胞活性无显著差异,转染48 h和72 h后, anti-miR-648组细胞活性较anti-miR-NC组显著下降;anti-miR-648组较anti-miR-NC组细胞的凋亡率明显上升,细胞和侵袭数目明显下降(P<0.05)。与anti-miR-648+si-NC组相比,anti-miR-648+si-Syk组中Cyclin D1、Bcl-2、MMP-2蛋白的表达量明显上升,Syk、P21、Bax、E-cadherin蛋白的表达量明显下降;转染24 h后,anti-miR-648组与anti-miR-NC组的细胞活性无显著差异,转染48 h和72 h后, anti-miR-648组细胞活性较anti-miR-NC组显著上升;anti-miR-648组较anti-miR-NC组细胞的凋亡率明显下降,细胞和侵袭数目明显上升(P<0.05)。综上所述,抑制Syk表达能够逆转抑制miR-648对胃癌细胞SGC-7901增殖、凋亡、迁移和侵袭的影响。
表4双荧光素酶报告实验
表5miR-648调控Syk的表达
注:与miR-NC组比较,aP<0.05;与anti-miR-NC组比较,bP<0.05
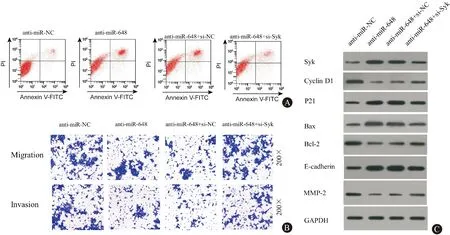
图5Syk及增殖、凋亡、迁移、侵袭蛋白的表达
表6抑制Syk表达能够逆转抑制miR-648对胃癌细胞SGC-7901增殖、凋亡的影响
注:与anti-miR-NC组比较,aP<0.05;与anti-miR-648+si-NC组比较,bP<0.05
表7抑制Syk表达能够逆转抑制miR-648对胃癌细胞SGC-7901迁移、侵袭的影响
注:与anti-miR-NC组比较,aP<0.05;与anti-miR-648+si-NC组比较,bP<0.05
3 讨论
胃癌是最常见的消化系统恶性肿瘤,其有高发病率、高死亡率、预后差的特点,严重威胁到人类健康[6]。目前手术根治是胃癌的主要治疗方法,早期患者术后5年生存率可达90%多,但因胃癌早期症状不明显及常规胃镜检查普及不足等原因,很大一部分患者在确诊时就已经处于胃癌进展期,错过了最佳手术治疗时间,5年生存率仅为20%左右[7]。现有胃癌治疗手段有限,单纯手术治疗的总生存率较低,放疗和化疗未能产生显著效果多用于术前术后辅助治疗,我国每年因胃癌去世的患者近352 300人[8]。因此,急需寻找到一种疗效好、不良反应小的新型治疗方法应对胃癌发病率和死亡率日益增长的现状。
大量研究表明,微小型RNA(microRNA,miRNA)是肿瘤发生发展中重要的参与者,已成为肿瘤研究中的热点。miRNA通过与靶基因的3′UTR完全互补切割靶基因的mRNA或不完全互补抑制其翻译,在转录后水平调控一系列重要生命过程[9],包括细胞的增殖、分化、凋亡、肿瘤细胞的浸润及转移等[10],在多种肿瘤中发挥作用,如张等人发现miR-648在前列腺癌中上调表达,并鉴定miR-648为新的候选前列腺癌miRNA生物标志物[11],Simone kreth 等人鉴定miR-648为胶质母细胞瘤中MGMT的重要转录后调节因子[12]。据研究报道,miR-648在胃癌细胞中表达上调[4-5],但其对胃癌细胞的具体生物学影响还未见报道。本课题通过RT-PCR法和Western blot法检测发现,与正常胃黏膜上皮细胞相比,胃癌细胞中miR-648的表达显著上调,与此前研究结论[4-5]一致,提示其可能起到促癌基因的作用。抑制miR-648的表达则细胞活性及迁移、侵袭数目显著下降,凋亡率显著升高,且Cyclin D1、Bcl-2、MMP-2蛋白的表达量明显下降,P21、Bax、E-cadherin蛋白的表达量显著上升。结果证明抑制miR-648的表达抑制胃癌的增殖、迁移和侵袭,促进其细胞的凋亡,但其具体分子机制尚不清楚。
本课题通过生物信息预测发现Syk可能是miR-648的靶基因, miR-648与Syk的3′UTR之间存在互补结合的核苷酸位点。脾酪氨酸激酶(Spleen Tyrosine Kinase,Syk)是非受体型蛋白酪氨酸激酶,在造血细胞、成纤维细胞、淋巴细胞、血管内皮细胞中广泛存在,在信号转导中发挥着重要作用[13]。近年研究表明,Syk的表达与肿瘤的发生发展密切相关,可以抑制多种恶性肿瘤的生长[14-16],如Syk可抑制乳腺癌生长和转移[17],抑制大肠癌大肠癌的浸润、淋巴结转移分化等[18]。 研究发现Syk在胃癌组织中下调表达,Syk的表达缺失与胃癌的生长和转移有一定的关联性[19-20],并有研究证明Syk对胃癌的生长转移有抑制作用,Syk可诱导胃癌组织中肿瘤细胞的凋亡[21]。荧光素酶基因报告实验和Western blot实验进一步证实miR-648可靶向调控Syk的表达,提示SKY是miR-648的下游靶基因。此外,研究还发现沉默Syk可逆转抑制miR-424对胃癌细胞增殖、凋亡、迁移和侵袭的影响,间接证实两者之间存在调控关系。
综上所述,本课题明确了miR-648对胃癌细胞增殖、凋亡、迁移和侵袭的影响,并初步探讨了miR-648可通过负调控Syk发挥抗肿瘤的作用机制,为胃癌的早期诊断和临床治疗提供了可靠的理论依据和新的治疗思路。
猜你喜欢
天津医科大学学报(2021年4期)2021-08-21 02:14:50
中日友好医院学报(2021年1期)2021-04-14 01:58:32
山东医药(2020年9期)2020-05-20 01:12:16
学苑创造·A版(2020年12期)2020-01-07 14:07:23
中国外汇(2019年15期)2019-10-14 01:00:34
作文教学研究(2016年1期)2016-07-05 12:22:47
法医学杂志(2015年4期)2016-01-06 12:36:36
法医学杂志(2015年4期)2016-01-06 12:36:36
医学研究杂志(2015年8期)2015-06-22 14:00:57
中国医药导报(2015年27期)2015-02-28 22:08:01
