
烟草低温诱导早花机理的研究进展
2019-11-22 03:08:52任雅萍朴世领
延边大学农学学报 2019年3期
任雅萍, 李 莹, 朴世领
(延边大学农学院,吉林 延吉 133002)
烟草是我国重要的经济作物[1],对经济发展起重要作用[2]。烟草早花是指烟株在生长过程中未达到当地栽培标准所需的株高及叶片数,过早开花现蕾、导致单株叶片数显著减少、造成大规模减产的现象[3]。事实上,这是处于恶劣环境条件下的烟株提前进入生殖生长阶段来繁衍后代保护种群的现象[4]。早花的烟株,植株矮小,叶片数锐减,叶片成分分布不均匀,使得烟叶品质参差不齐,经济效益大幅度降低[5-7]。
除烟草本身的品种特性外,诱导烟草早花的主要环境因素为光周期与温度[7]。由于我国北方烟区早春气候不稳定,烟株易受到天气低温等不良因素的影响,从而造成早花[8-9]。目前国内对烟草早花的研究,主要针对于早花发生原因和预防补救措施等方面,而关于耐低温早花分子机理方面的研究少之又少。防止早花的根本途径是挖掘耐低温早花基因、培育耐低温抗早花的烟草新品种,从根源上预防早花的发生。
1 低温诱导下早花烟株的生理生化变化
温度是影响植物生命活动正常进行的主要因素,对烟叶的品质提高有很大的作用,烟草喜温不耐寒,温度大幅降低并且持续10 d以上就可能引起早花[10]。因此,温度的高低在烟草的生长发育过程中发挥了重要作用。王欣亚等[11]通过调查低温(12 ℃)处理下的吉烟9号与云烟87号的保护酶(SOD、POD、CAT)活性发现,处理组与对照组相比,保护酶活性均呈现出先升后降的趋势,但始终低于对照组,且对照组保护酶活性呈平稳上升趋势,处理组现蕾率提前且现蕾率高于对照组。王浏杰等[12]以NC82和K326进行苗期低温(14 ℃)处理后发现,脯氨酸(Pro)和丙二醛(MDA)含量均呈现出先增高后降低的趋势。上述结论同时也说明,虽然烟株在受到低温胁迫后会通过产生保护酶、Pro等物质维持细胞内动态平衡,但这种调节效率不高,不能很好地防止早花的发生,通过由细胞膜受损后的产物MDA的含量也可以看出,低温对于烟株早期的影响较大,随叶龄的增加,这种不利影响有所缓解,但不可否认这种影响会导致烟株生育期提前,影响烟株的成花调控最终影响成花基因的表达。低温引起的早花不仅对烟株的生长发育产生众多不利影响,对烟叶本身的质量也有着巨大影响。以烤烟品种K326进行低温处理试验表明,低温处理过的叶片中可溶性糖含量升高,蔗糖磷酸合成酶SPS活性升高,淀粉酶活性降低,淀粉在叶脉处积累增加[13],直接影响了烟叶的品质。以上研究内容直接说明了低温处理后烟株由于生殖生长增强而降低烟叶产量的同时,促进了次生代谢物的产生和积累,导致烟叶碱和氯含量增加,增加了糖类的分解和消耗,从而影响了烟叶内部化学成分的比例,直接引起烟叶商品价值降低从而增加经济损失。
2 烟草成花花熟状态期的研究
自然条件下,植物生长到一定时期会具有感受外界环境条件的能力,称之为花熟状态[14]。进入花熟状态的植株,当其接受所需环境信号的诱导,特别是温度和光周期的诱导后,分生组织进入一个相对稳定的状态,称为成花决定态。进入成花决定态的植物细胞,已经具备了分化为花的能力但还未分化[15]。而植物是否进行花的分化,与植物分生组织处于成花决定态的细胞数量有关,达到一定数量,就可以启动花的发生,植物开始进入花的发育,而未达到一定数量时则发育为叶和节[16]。
在烟草的成花决定机理研究中,依据植株是否具备感受外界信号的能力从而影响其生长发育提出2种理论:1) 在正常的外部生长条件下,烟草的花期取决于自身的遗传特性[17],并且不同种类烟草(长日、短日、日中)在花芽分化时已经具有相对一致并且可预测的节数以及节间数[18-19]。Mcdaniel等[19]指出,烟草在23~36片叶龄时,顶端分生组织开始由营养分生组织转入花芽分生组织。Susan等[20]研究发现,烟草(Wisconsin38)20片叶龄前顶芽还未进入成花决定态,21片叶则开始进入成花决定态,大约需2 d后完成顶端分生组织的成花决定,而顶芽进入成花决定态后,还会继续产生4个节,即25片叶时,顶花开始发育。2) 烟草的成花是由营养生长阶段至生殖生长阶段逐步进行的[21]。对于烟草来说,在后期的可变营养生长阶段,生长锥分化为叶芽还是花芽与环境条件有关。而当环境条件适宜花芽分化时,就逐渐转向花芽分化[22]。有研究者认为,烟草生长到6~12片叶时是其低温敏感阶段[23];也有研究表明,白肋烟8片叶龄前后是对低温和短日照的敏感时期[23]。
由此可见,烟苗在一定的生长阶段,即具有一定数量的叶片时才能够感受外界信号的刺激,进入成花决定态,但并非所有叶片都具备了感受外界刺激的能力,而是达到一定叶龄的叶片才能感受到低温或者光周期,从而实现花芽的分化[24]。所以植物细胞如何感受外界环境诱导,如何过渡到成花决定态,以及如何启动花发育基因还需进一步的研究。
3 烟草低温诱导早花分子机理
3.1 调控植物开花途径的重要基因
成花诱导是高等植物从营养生长转入生殖生长的关键步骤,其过程十分复杂,同时受植物内在因素和外界环境条件的影响,而环境条件作为相对可控的因素,研究其作用方式及形成机理,对植物的成花研究有十分重要的意义。目前对植物成花的分子研究主要分为光信号,低温诱导成花以及特异基因的表达3个方面[25]。遗传学分析已经证明主要有6种成花途径存在于拟南芥中[26],即春化途径、自主开花途径、光周期途径、赤霉素途径、温敏途径以及年龄途径。这些途径都通过调控开花途径的关键基因FLOWERINGLOCUST(FT)、SUPPRESSOROFOVEREXPRESSIONOFCO1(SOC1)和LEAFY(LFY)来控制开花的确切时间[27]。这些基因又受2个中枢上游调控子(central upstream regulator)CONSTANTS(CO)和FLOWERINGLOCUSC(FLC)的逆向(antagonistically )调控[28]。作为光周期的正调控子(positive regulator),CO可以编码1个调控转录的锌指蛋白,此表达不仅受生理因素和光周期的控制,转录后水平的稳定性也受光周期的控制[28-30],FLC[31]编码1个MADS- box转录因子,介导自主开花途径以及春化途径,是开花的负调控子(negative regulator),具体开花途径及其关键基因见表1。

表1 调控拟南芥开花的主要途径及其关键基因
3.2 低温影响烟草开花的相关基因
温度诱导下的成花作用也就是春化作用,它能够克服光周期的阻碍,促进植物开花。目前,关于烟草耐低温相关基因的研究较少,而拟南芥是目前作为在成花方面被人们研究的最为透彻的植物,其成花机制的研究表明[31-33],由低温诱导成花的调控基因FRIGIDA(FRI)、FLOWERINGLOCUSC(FLC)、VERNALIZATION1(VRN1)、VERNALIZATION2(VRN2)和VERNALIZATIONINSENSITIVE3(VI-N3)通过环境条件的改变和一系列生化反应后,进一步调控开花途径的关键基因FT、SOC1和LFY来控制开花的确切时间。
3.2.1FLC基因
低温诱导植物提前开花的现象主要由几个MADS-box转录基因控制,其中最重要的是FLC基因[30-32]。FLC基因作为开花的负调控子,在植物的所有部位均有所表达,受多种成花基因的调控,具体调控途径见图1。在低温条件下,FLC基因的表达受到抑制,从而使芽中SOC1等相关基因表达,诱导植株开花[34],造成早花。通过进一步对拟南芥自然生态型的基因分析表明,植物对春化作用的反应是由2个显性基因FRI和FLC的协同增效作用所决定[32-33],FRI编码的蛋白,可使FLC的mRNA转录水平升高[34]。也就是说,当FRI基因有活性时,FLC基因的转录水平高且稳定,植物表现出晚花。
Sheldon等[35]用含有FLC反义基因的T2C24-转基因系进行试验,在花椰菜病毒的35S启动子的控制下降低FLC基因转录水平,发现植物出现早花现象,但其对春化作用无响应。进一步的研究发现,随着低温处理时间的延长,FLC基因的转录水平越来越低,直至几乎检测不到。李元元等[36]以低温敏感品种NC82为试材,通过同源克隆方法获得了NtFLC全长序列,进一步用半定量RT-PCR技术对NtFLC在不同组织表达进行定性分析结果表明,该基因可能与NC82低温敏感、易早花特性相关。
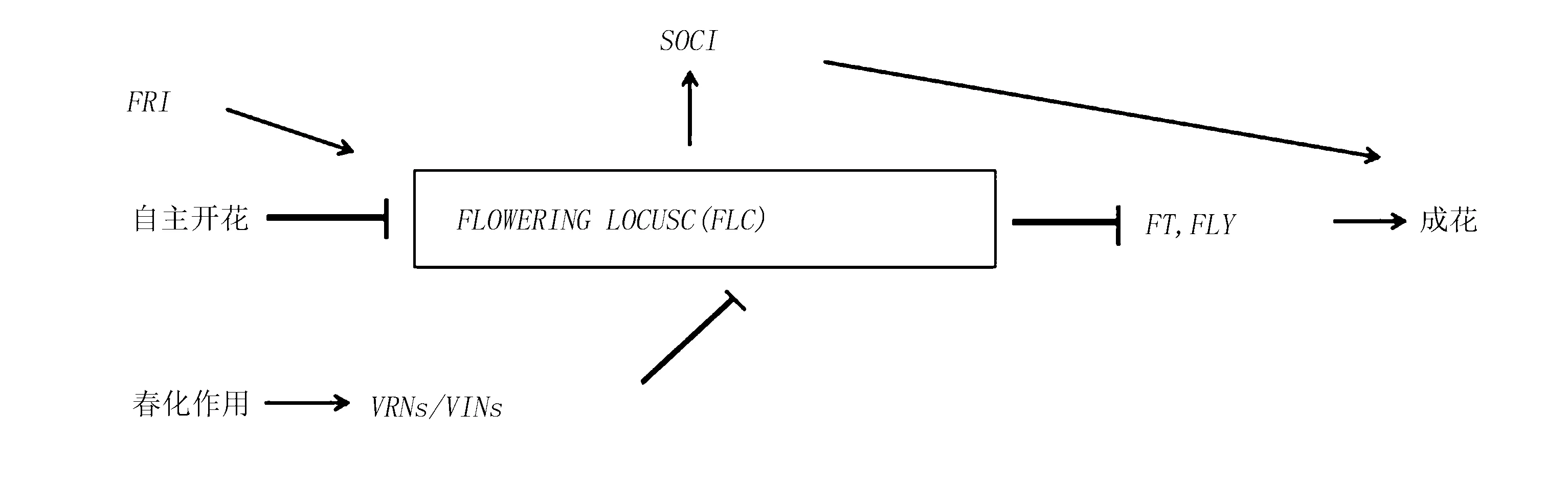
注:图中箭头表示促进基因的表达;直线表示抑制基因的表达。
图1FLC基因拮抗调控表达的不同途径(Marquardt S等,2014)
Fig.1 Different pathways ofFLCgene antagonistic regulation and expression(Marquardt S,et al,2014)
3.2.2FRI与VRN基因
Gazzani S[37]通过对拟南芥的研究发现,FRI基因的等位变异基因被认为是影响拟南芥春化过程和开花时间的主要决定因素。FRI基因的作用是增加成花抑制因子FLC基因的RNA水平,而FLC基因抑制了向开花过渡所需基因的表达量。在春化过程即低温条件下,FLC基因的RNA水平降低。Johanson等[38]对5个早花品种拟南芥的分子基础进行分析,发现不同实验材料的FRI等位基因均有不同程度变异或功能缺失,因此认为拟南芥开花时间的变化可能是通过产生无功能或功能弱化的FRI,进而减弱了对FLC表达量的促进作用引起的。
VRN基因也是重要的春化基因之一,目前关于烟草VRN基因的研究鲜有报道,但其在冬小麦中的研究表明,VRN1基因是显性基因,其转录水平随春化作用的进行逐渐上调,而VRN2基因的转录水平随春化作用的进行逐渐下调[39]。Loukoianov 等[40]研究发现,VRN1基因在二倍体和多倍体小麦的早期发育阶段起重要作用,通过RNA干扰减少VRN1基因转录水平,推迟了小麦生长点向生殖阶段的过渡,增加了叶片数量,并推迟了2~3周的抽穗时间。也有研究认为VRN基因中的VRN2、VRN1与FLC稳定的低水平表达有关[40-41]。Levy等[42]研究认为,拟南芥在长日条件下,VRN1等位基因的突变体vrn1-1、vrn1-2没有延迟开花,但对春化作用的响应性有所降低,在供试的变异植株fca-1和vrn1-1 fac-1中,6周的低温处理均可降低FLCmRNA的表达水平。但低温处理后的vrn1-1 fac-1植株转入正常温度条件一段时间后,FLC基因的表达水平又重新升高。这些结论说明,VRN1、VRN2虽然不是低温负调控FLC基因表达所必需的因子,却在维持在后期成花温度条件下FLC低水平稳定表达中起关键性作用。
3.2.3 低温影响下与信号通路有关的成花基因
在拟南芥中关于花芽分化中主效基因FLC的表达,也受到一类甾醇类植物激素—油菜素内酯(BR)信号通路的调控,BR信号可以调控通路衰弱促进FLC基因的表达,使植物花芽分化延迟,从而影响早花现象的发生。肖立增[43]研究发现,在云烟 85和兴烟1号苗期低温处理时,与苗期低温相应有关的主要基因NtBRI1以及涉及信号通路中的 Nt BSK1、Nt BSK2和NtBSK3基因的表达量明显高于常温处理,且荧光定量PCR检测结果表明,NtBRI1随着低温处理时间持续,这种受体表达的差异明显升高。进而以兴烟1号为实验材料的研究表明,低温诱导使NtFLC的表达量受到了抑制,其表达量降低,对植株成花的抑制效果也减弱,造成花芽分化提前,但是这种抑制效果的强弱不与BR信号转导通路运行活跃程度的强弱存在绝对的正相关关系,由此说明BR信号转导途径在调控植株花芽分化时间上受春化作用的影响较小,可能存在另一条独立的调控途径。
赤霉素途径也是开花过程中能非常重要的途径,赤霉素(GA)是调控植物生长发育的一种重要激素。在拟南芥种子萌发过程中受到低温胁迫后,种子中特定GA生物合成路径中负责催化的活性GA(GA1、GA4)的合成酶(GA20-oxidase, GA3-oxidase)的编码基因表达量上升;而活性GA代谢途径中负责催化产生非活性GA的酶(GA2-oxidase)的编码基因表达量下降[44];即活性GA(GA1,GA4)含量升高,非活性GA含量降低,推测低温胁迫也可能会诱导植物其它组织器官中的活性GA含量升高。在拟南芥中的研究表明[45],At-MYB33可以受赤霉素诱导,使其在茎尖有较高水平转录,并且AtMYB33能够结合LEAFY基因的启动子区域中保守的8碱的Motif,说明AtMYB33可能通过GA信号调控植物开花。白戈等[46]研究根据烟草芯片数据和PCR扩增技术克隆到一个在进化过程中高度保守的MYB类启动因子基因,命名为NtMYB15基因,将该基因转入烟草K326中,低温胁迫的转基因植株与低温胁迫的非转基因植株对比,表现出相对晚花的特征,说明NtMYB15在栽培烟草中参与低温胁迫诱导开花调控。但NtMYB15基因参与调控烟草开花时期的机制目前并不清楚,且NtMYB15基因可能参与到GA调控烟草成花过程,但需要进一步实验验证此推论。
茉莉酸(Jasmonic acid,JA)及茉莉酸甲酯(Methyl Jasmonate,MeJA)等衍生物,统称为茉莉素(jasmonates,JAs),是一种近年来发现的植物激素,广泛存在于各种高等植物中。目前,在烟草中已经克隆了JA信号通路中的关键基因NtAOC、NtAOS、NtCOI1、NtJAZ1、NtLOX、NtMYC2[47],冉燕子等[47]通过克隆在JA信号通路中扮演重要角色的NtNINJA基因,发现该基因协助JAZ蛋白抑制JA信号转导,进一步测量低温处理烟草中的JA含量发现,JA含量均在低温处理开始后迅速上升,直到低温处理结束后才逐渐下降,表明低温胁迫增强了烟草JA信号通路的活跃性,合成大量JA提高对低温胁迫的抗性。但对于防治烟草早花方面来说,在低温胁迫条件下,茉莉酸调控烟草开花的关系尚不明确,有待进一步研究。
3.2.4 低温条件下影响烟草成花其他相关分子研究
杨永银等[48]研究发现,低温会诱导烟草茎顶端分生组织中抗坏血酸氧化酶(AO)的活性,表达水平增加,脱氢抗坏血酸(DHA)含量增加,且DHA能诱导开花关键基因SOC1表达水平升高;将AO基因克隆,通过农杆菌介导的转化获得AO转基因植株,并筛选得到AO过表达植株,与野生型相比,AO过表达植株出现植株矮化,生长缓慢,开花提前,气孔较大但密度低等现象,且光合速率明显高于野生型。
miRNA是与靶mRNA的互补配对的非编码小RNA,可以在转录后水平上对基因的表达进行负调控,导致mRNA的降解或翻译抑制[49]。其中与开花时间提前有关的主要为miR156和miR172[49],转入miR156“模拟序列”的植株,由于该序列的过表达导致植株缺少幼年期而提前开花[50]。也有研究表明,miR172的过表达能让拟南芥开花提前[51]。然而目前有关miRNA调控植物花发育进程的研究主要集中在拟南芥、金鱼草、水稻、玉米等植物上[52],在烟草上未见报道,但这些研究或许可以为烟草早花分子机理的揭示提供借鉴,有利于全面的探究烟草成花分子机理以及生长发育过程。
4 展望
早花问题一直制约着烟草产业的发展。不同烟草品种遗传特性不同,受低温等外界环境因素影响开花时间的程度也有所不同。在实际生产中,当早花发生时则采取一些农艺措施来解决烟草植株叶数锐减的问题,但这是一种极为耗费人力物力的补救措施,也无法彻底解决早花的问题。而通过对烟草耐低温诱导早花主效基因的发掘,利用分子标记辅助选择、转基因技术以及分子设计育种技术等选育出耐早花的烟草品种,能够从根本上解决烟草发生早花的问题,有利于最终切实提高烟草的产量和质量。
目前分子生物学正快速发展,但从分子角度解释调控烟草开花的机理及其相关分子表达的研究严重不足,尤其是低温诱导烟草早花分子机理的研究更是少之又少,例如烟草的低温诱导成花途径究竟有哪些?又有哪些关键基因参与了成花的调控?在低温条件下,各个途径如何相互制约和相互促进导致烟草提前开花,相应关键基因的表达发生何种变化?miRNA在成花过程中究竟起多大作用等一系列问题还有待于进一步地研究和探讨,缺乏一个系统而全面的理论体系。
猜你喜欢
中学生天地(A版)(2023年1期)2023-02-17 00:33:04
蔬菜(2022年1期)2022-11-11 01:08:01
麦类作物学报(2022年4期)2022-05-23 03:29:20
金山(2021年10期)2021-11-02 09:11:52
热带生物学报(2019年1期)2019-04-18 01:54:04
江苏农业科学(2018年11期)2018-06-29 05:06:20
生命科学研究(2018年1期)2018-05-29 01:12:47
现代园艺(2017年23期)2018-01-18 06:57:46
上海农业学报(2017年3期)2017-04-10 12:39:22
上海农业学报(2017年3期)2017-04-10 12:39:14
