
不同密度樟树(Cinnamomum camphora)幼苗生长和叶片性状对氮磷添加的响应
2019-11-07 06:43:36林婉奇蔡金桓
生态学报 2019年18期
林婉奇,蔡金桓,薛 立
华南农业大学林学与风景园林学院, 广州 510642
工业革命以来,人类活动(如化石燃料燃烧,森林干扰和土地变换等)改变了氮素从大气向陆地生态系统输入的方式和速率,进而导致森林生态系统养分变化失衡[1,2]。土壤中N和P的添加均能影响树木的生理代谢过程。氮素作为植物生长的必需元素,是构成植物各种光合酶、ATP和NADP+的重要成分[3]。施N导致叶片氮含量的增加,从而促进了植物叶绿素浓度的升高以及生物量和光合碳的积累[4]。相关研究指出,N沉降增加了植物可利用N的含量,有助于植物的生长发育[2,5,6]。
植物的生长主要由养分的吸收与利用决定,N和P的吸收、同化和转运直接影响植物的生长发育[7]。目前,国内外关于氮沉降对植物生长的影响已有较多研究。一般而言,N添加在一定范围内能够促进植物的生长[8,9],但是长期过量的N添加会抑制植物的生长速率,严重时还会导致植物死亡以及生态系统的衰退[10]。Wang等[6]指出,幼苗生长主要取决于N沉降的速率。与N添加相比,P添加在这方面的研究相对较少。杨晓霞等[11]在研究氮磷添加对青藏高原高寒草甸植物的影响时发现,N和P均促进了禾草的生长,但是P添加的促进作用更大。
种植密度是林分竞争的重要研究内容,影响着植物对环境资源的利用和所占有空间的获取[12]。Xue等[13]报道密度影响尾叶桉的胸径生长,但是对树高生长的影响不明显。宿爱芝等[14]的研究发现辣木(Moringaoleifera)的株高、地径和冠幅与种植密度之间表现出负相关关系。朱仕明等[15]的研究也表明,高密度会抑制乐昌含笑(Micheliachapensis)的苗高和地径的生长,这主要是个体植株间对光照、水分和养分等资源竞争的结果[16-17]。但是,肖玲玲等[18]、黎磊和周道玮[19]的研究却表明,高密度种植促进了樟树(Cinnamomumcamphora)和红葱(Alliumcepavar.proliferum)株高的生长。
樟树(Cinnamomumcamphora)作为樟科樟属常绿乔木中经济价值最大的树种之一,是我国特有的珍贵用材、芳香油类以及城镇园林绿化树种。有关学者对樟树的生长和光合[20]、土壤氮矿化速率[21]、土壤呼吸[22]、凋落物及养分动态[23]、生长和重量分配格局[8]、繁殖方法[24]以及逆境胁迫[25,26]等进行了研究,但是尚未见到氮磷添加和种植密度的交互作用对樟树幼苗生长影响的报道。本研究选取樟树幼苗作为实验对象,对其生长形态进行了测定,旨在了解不同种植密度樟树幼苗对氮磷添加的响应机制,以期为全球化背景下樟树人工林生态系统的经营管理提供依据。
1 材料与方法
1.1 研究区概况
试验地位于广州市华南农业大学板栗园(113°21′E,23°09′N),全年平均气温为21.9℃,年降雨量约为1736 mm,集中在4至6月,平均相对湿度77%,水热同期,雨量充沛,属亚热带季风气候。土壤为花岗岩发育而成的赤红壤。试验地环境光线充足,适合幼苗的生长。
1.2 试验设计
以广东省国森苗圃所提供的樟树1年生实生苗为试验材料,幼苗的平均苗高为46.7±3.38 cm,平均地径为0.36±0.07 cm。采用直径35 cm,深30 cm的无纺布美植袋种植,基质为该试验地0—20 cm土层的土壤。
根据样地的氮沉降水平背景值以及参考同类研究方法[27,28],大气氮沉降中近3/4为铵态氮,本试验选择氯化铵(NH4Cl)模拟大气氮沉降,并且以二水合磷酸二氢钠(NaH2PO4·2H2O)模拟大气磷沉降。
试验时间为2017年6月至9月。试验采用4×4双因素析因设计16种组合(表1)。氮磷沉降处理设置4个水平:不施肥(CK)、施N肥、施P肥和施N+P肥。N、P和N+P施肥量分别为40 g m-2a-1(NH4Cl)、20 g m-2a-1(NaH2PO42H2O)和40 g m-2a-1(NH4Cl)+ 20 g m-2a-1(NaH2PO4·2H2O)。种植密度设置4个水平:10、20、40和80株/m2(分别用密度I、II、III和IV表示),即在直径35 cm的美植袋中分别种植均匀1、2、4和8株幼苗,尽量保持每盆内的幼苗距离相近。N、P和N+P施肥量分别为NH4Cl 3.84 g 袋-1a-1、NaH2PO4·2H2O 1.92 g 袋-1a-1和NH4Cl 3.84 g 袋-1a-1+NaH2PO4·2H2O 1.92 g 袋-1a-1。试验采用完全随机区组设计,4个密度和4种施肥处理交叉共产生16个处理,每个处理3次重复,合计48个小区,每小区16株幼苗。根据处理水平的要求,模拟氮磷沉降时,将各处理每次所需质量的肥料溶于12升的水中,向每盆的土壤浇灌200 mL溶液,保证不会引起烧苗的现象,不施肥处理的只施等量的水。自6月起每月月初和中旬分2次向幼苗施肥直至收获,共施肥8次。
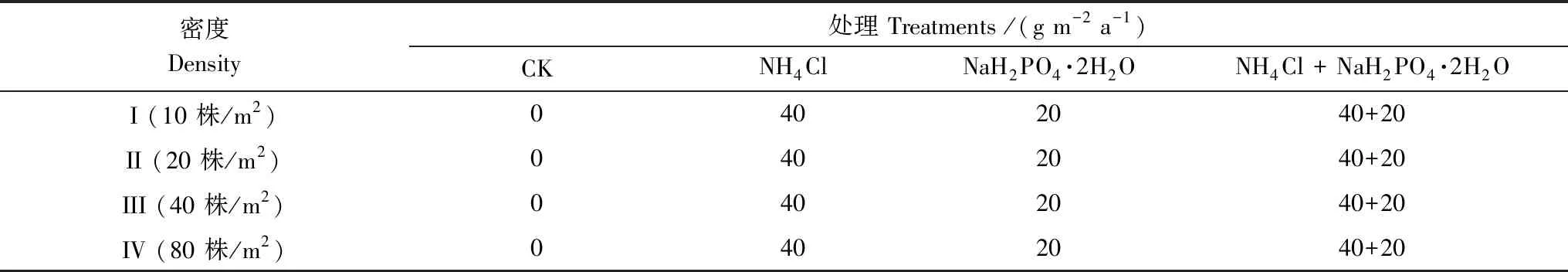
表1 樟树幼苗氮磷和密度处理水平
1.3 地径和苗高测定
在2017年5月初测定幼苗的地径和苗高,作为初始值。在2017年9月底再测定一次幼苗的地径和苗高态。每个处理随机选取10株长势一样的幼苗,每株幼苗选取30片健康无病虫害的叶片,采用便携式叶绿素SPAD仪测定各处理叶片的叶绿素相对含量,并测量其叶面积,以便求出比叶面积SLA(比叶面积=叶面积/叶干重)。
1.4 数据处理
用SAS 9.3统计分析软件对生长形态进行Duncan多重比较,Microsoft Excel 2003对数据进行平均值、标准偏差分析和作图。
2 结果与分析
2.1 氮磷添加下不同种植密度樟树幼苗的生长状况
2.1.1平均单株幼苗苗高
N和N+P处理的各密度幼苗的苗高均大于CK,在密度I和II幼苗中,P处理的幼苗苗高小于CK,而密度III和IV的幼苗苗高相反。密度III幼苗的N+P处理苗高显著大于CK(P<0.05)(图1)。随着密度的增大,CK、N和P处理的樟树幼苗苗高均呈现下降的趋势,除了P处理外,其余处理的苗高显著减小(P<0.05),N+P处理的变化规律不明显。

图1 不同处理和密度的樟树幼苗的苗高和地径Fig. 1 Seedling height and Ground diameter of Cinnamomum camphora seedlings under different treatments and densities同一测定内容用不同小写字母表示0.05水平上的显著差异
2.1.2平均单株幼苗地径
与CK相比,密度I、II和III幼苗中,N和N+P处理均能促进樟树幼苗地径的增长,密度I和II幼苗中N+P处理的地径显著增加(P<0.05)。密度IV幼苗中N和N+P处理的均抑制了幼苗地径的增长;密度I和II幼苗中,P处理促进幼苗地径的增长,密度III和IV的幼苗则相反。随着密度的增大,N、P和N+P处理的樟树幼苗地径均显著下降(P<0.05)(图1)。
2.2 氮磷添加下不同种植密度樟树幼苗叶片性状
2.2.1幼苗叶片的SPAD值
与CK相比,除了密度II幼苗的P处理外,N、P和N+P均增大了各密度的幼苗叶片的SPAD值,其中密度I和V的三种处理、密度II和III的N+P处理达到显著水平(P<0.05)(图2)。随着种植密度的增大,CK、N、P和N+P处理的幼苗叶片SPAD值均呈现下降的趋势,多达到显著差异(P<0.05)。

图2 不同处理和密度的樟树幼苗的SPAD值和比叶面积Fig.2 SPAD and SLA of Cinnamomum camphora seedlings under different treatments and densities
2.2.2不同处理下幼苗叶片的比叶面积SLA
与CK相比,密度I和II幼苗中N和N+P处理的幼苗SLA增加,密度I、II和III幼苗的P处理的幼苗SLA减少。密度III幼苗各处理的SLA减少,而密度IV幼苗各处理的SLA增加,但除了密度I和II幼苗的P处理外,各处理的幼苗SLA差异不显著(图2)。随着种植密度的增加,各施肥处理下樟树幼苗的SLA变化规律不明显,但在密度IV幼苗中各处理的SLA均显著大于其他密度相同处理的幼苗(P<0.05)。
2.3 氮磷添加与密度对樟树幼苗生长和叶片性状的交互作用
施肥和密度处理对樟树幼苗的苗高、地径和SPAD值均有显著影响(P<0.05),对SLA没有显著影响,密度和氮磷添加对叶片SPAD值产生显著的交互作用(P<0.05)(表2)。

表2 不同密度和施肥处理对樟树幼苗生长和叶片性状的交互作用
F值是组间与组内的离差平方和与自由度的比值;F值越大说明处理之间的效果越明显
3 讨论
3.1 氮磷添加对不同种植密度樟树幼苗生长状况的影响
幼苗是植物生长历程中最敏感的阶段,其生长指标在适应环境因子的变化过程中能表现出一定的可塑性,对于研究植物在氮磷沉降等养分环境下的适应机制具有指示性[29]。本研究结果显示,N、P和N+P处理对樟树平均单株幼苗的苗高和地径整体上有促进作用。N作为植物生长阶段中最重要的养分限制元素,可以通过影响植物体内叶绿素和可溶性蛋白含量以及光合酶的合成与活性,进而引起光合作用的变化,最终导致植物的生长发育和形态构建发生改变[30,31]。适量的N输入通过降低硝化菌、反硝化菌和植物对矿质氮的竞争,增加土壤N的有效性,从而促进植物的生长[32]。其他氮添加实验也有相似的结果,如吴茜等[30]还发现,施氮显著促进了秃瓣杜英(Elaeocarpusglabripetalus)、枫香(Liquidambarformosana)和木荷(Schimasuperba)的苗高和地径生长,但是对青冈(Quercusglauca)和马尾松(Pinusmassoniana)的苗高促进作用却不显著。这可能是因为不同树种对N的利用和分配不同,因而对氮添加的响应也不相同[9]。肖迪等[8]研究三个种源的五角枫(Acermono)幼苗对氮添加的响应时发现,苗高和地径在中高氮水平下达到最大值。王晓荣等[33]的研究发现,氮添加亦能促进麻栎(Quercusacutissima)和栓皮栎(Quercusvariabilis)的株高和地径增长。
本研究中,P处理对幼苗生长的促进效果较弱,N+P处理下的效果最好。这可能是因为土壤中的Ca、Fe、Al和土壤粘粒等通过沉淀、吸附、固定以及微生物吸收作用,将有效P转化为 Al-P、Fe-P、Ca2-P和Ca8-P等闭合形态的难溶性磷,没有大幅增加P的有效性,故对幼苗的促进作用相对较弱[34]。Elser等[35]和Harpole[36]等的研究也发现,N和P共同添加对植物生长的促进效果优于单一添加,这是因为植物光合作用需要大量的酶(主要为Rubisco酶,N为其主要组分) 参与,而这类酶由核糖体(P为其主要组分)合成。Agren等[37]指出,植物体内N和P在吸收和利用上相互依赖,并且P添加有利于促进生态系统中N利用效率的提高[36,38],且本试验地为南方红壤地区,普遍缺磷,氮磷添加共同作用下给土壤大量输入了N和P,使幼苗能够充分吸收养分,并把多余的N分配给磷酸酶以增强其活性,同时缓解P限制,故长势优于单一的P处理。N和P对植物生理生态特性的影响还与树种类型、群落组成和土壤养分本底有关[39]。刘双娥等[40]的研究发现N添加促进了马占相思、木荷和马尾松幼苗的生长。庞丽等[41]的研究表明,N添加增加了马尾松和木荷的地上部分生长量,严重抑制了杉木幼苗的生长。庞丽等[42]的研究发现,氮处理增加了马尾松根系有机酸和酸性磷酸酶的分泌, 使植株极度缺磷, 抑制了幼苗的生长。
本结果表明,随着种植密度的增加,樟树平均单株幼苗的苗高和地径均呈现下降的趋势。李洁等[12]研究不同种植密度对银合欢(Leucaenaleucocephala)生长的影响发现,高密度种植下银合欢的侧枝生长空间小,枝叶重叠,邻体干扰强,叶片的光合速率减弱,抑制了树木的生长[43,44]。朱仕明等[15]的研究也发现,高种植密度抑制了乐昌含笑(Micheliachapensis)的地径和苗高的增长。因为种植密度会影响种群内个体可获取的资源量,种植密度越大,植物间距离越小,相邻间植物对有限的光照、水分和养分的竞争愈加激烈,因而幼苗的生长受抑制。本研究中,在高种植密度下,N+P处理下幼苗的生长状况优于CK处理。这可能是因为氮磷添加中N和P的大量输入缓解了高种植密度下幼苗间对养分的竞争程度。
3.2 氮磷添加对不同种植密度樟树幼苗SPAD和SLA的影响
外源性N或P的输入对植物叶片的生长和生理机能的影响主要体现在促进或抑制叶片生长、改变叶绿素含量从而促进或抑制光合速率以及改变叶片形态结构和养分状况[45]。叶片是植物进行生理活动和发挥生态功能作用的主要器官[17]。叶片中光合色素含量是反映植物光合能力的一个重要参数,光合色素中的叶绿素对光能的吸收、传递、利用和光化学反应具有重要作用[46]。叶片SPAD值表示的是叶绿素的相对含量,与叶绿素含量呈正相关关系,在一定程度上能够反映植物叶绿素含量的变化[47,48]。本研究结果表明,与CK处理相比,N、P和N+P处理在整体上促进了幼苗叶片SPAD值的上升,其中N和N+P处理的效果更好, 这是因为N是构成叶绿素的主要成分,光合产物的运输离不开P的参与,叶中的氮和磷的含量与光合速率密切相关,氮磷添加处理有利于改善土壤养分状况,促进叶片叶绿素含量的增加。随着密度的减少,个体的空间逐渐增大,幼苗对N、P及其他叶绿素合成元素的吸收量增加,最终导致叶绿素含量水平的提高,因此氮磷添加和低密度对叶片SPAD值产生显著的交互作用[49]。
3.3 氮磷添加和种植密度樟树幼苗的交互作用
本研究中,氮磷添加对幼苗高生长和地径所长具有显著正效应, 密度则具有显著负效应,而氮磷添和密度的交互作用对幼苗生长没有显著影响,说明密度消除了氮磷添加对幼苗生长的正效应。氮磷添加促进了根系生长发育,而增加的种植密度减少了幼苗根系生长发育的空间,抑制了幼苗细根生长,消除了因土壤养分增加而促进根系生长发育的正效应。叶片的SPAD可以反映叶片的光合能力。氮磷添加、密度及氮磷添加和密度的交互作用对叶片SPAD均产生显著影响。尽管氮磷添加对幼苗具有显著正效应, 但是密度为显著负效应,前者正效应大于后者的负效应,所以氮磷添加和密度的交互作用仍然显著促进了SPAD的增加。
猜你喜欢
今日农业(2022年14期)2022-11-10 00:27:51
新作文·小学低年级版(2022年4期)2022-08-30 07:23:00
阅读(科学探秘)(2020年8期)2020-11-06 06:22:48
北方音乐(2019年6期)2019-07-16 07:50:53
中国果业信息(2019年1期)2019-01-05 17:41:42
现代园艺(2018年3期)2018-02-10 05:18:14
生物学教学(2017年9期)2017-08-20 13:22:32
小哥白尼·趣味科学画报(2014年9期)2015-04-07 21:01:12
食品工业科技(2014年6期)2014-05-10 06:04:50
天然产物研究与开发(2014年6期)2014-04-27 14:15:59
