
毛白杨树干呼吸及其温度敏感性季节变化特征
2019-11-07 06:43郭丽丽杨庆朋乔雅君张茜茜梁伟佳郑云普
生态学报 2019年18期
刘 亮,李 菲,郭丽丽,杨庆朋,乔雅君,聂 唯,杨 彬,张茜茜,曹 旭,梁伟佳,郑云普,*
1 河北工程大学水利水电学院, 邯郸 056038 2 中国科学院沈阳应用生态研究所/中国科学院森林生态与管理重点实验室,沈阳 110164 3 河北雄安新区生态环境局,雄安 071700 4 中国农业大学水利与土木工程学院,北京 100083 5 中国水利水电科学研究院水利研究所,北京 100048
森林自养呼吸过程是陆地生态系统碳循环的重要组成部分[1],已经逐渐成为当前全球变化生态学研究领域的热点问题之一[2,3]。以往的研究结果表明,森林生态系统总初级生产力的50%—70%通过自养呼吸的形式进入大气[4- 6]。同时,树干呼吸作为森林生态系统碳循环过程的重要组成部分,约占整个森林生态系统自养呼吸的12%—42%[7]。尽管如此,树干呼吸相关研究却在整个森林生态系统碳循环研究中被关注的相对较少,以至于目前对于森林生态系统碳储量的估算还存在很大的不确定性[4,6]。
树干呼吸(Es)是一个非常复杂的生物学过程,CO2气体主要是由树干木质部、韧皮部和形成层等组织的活细胞通过生理的有氧呼吸代谢过程而产生[5]。树干呼吸过程不仅受到温度、水分和光辐射等多种环境因素的影响[2,7],而且还会根据树干液流、生长速率和林分状况的不同而发生明显变化[1,3,6]。因此,难以准确判断究竟哪些关键因素直接决定着树干的呼吸过程。通常而言,温度作为重要的环境因素可能直接影响Es[8- 10]。目前,大多数学者认为温度可能通过多个不同的途径对树干呼吸产生影响,主要包括温度升高增加细胞的呼吸速率,增加CO2的扩散系数或者降低液流中溶解CO2的浓度[10-14]。以往许多研究已经发现Es和树干温度之间具有较好的相关性[11],故研究者往往利用以温度为变量的函数估算Es,且认为Es随着树干温度呈现指数增加的趋势,其与树干温度的关系一般表示为温度敏感指数(Q10)[12- 15]。然而,另外的一些相关研究却发现Es和树干温度的相关性并不显著,甚至根本不存在相关性[9,16],即使将Es进行温度标准化校正后,Es在树种和林分间却仍存在较大的时空变率[17]。因此,树干Es及其对温度的响应(Q10)呈现出较大的时空变异性,以至于对森林生态系统碳储量的估算造成偏差[3,6],这在一定程度上限制了森林碳平衡模型的发展[4]。
以往大多数的研究结果表明,树干呼吸及其温度的敏感性随季节的不同而呈现出明显的动态变化特征[10,13,16]。在不同季节里林木的光照条件、土壤水分状况、氮素利用率等都可能存在较大的差异[15,17],从而导致树干内部的非结构性碳水化合物含量的改变[12],最终造成树干Es和Q10的显著变异[16,17]。杨庆朋[18]对我国亚热带典型人工林连续2年的监测数据发现,3个树种生长季的Q10值均明显低于非生长季,且将生长季和非生长季的数据整合一起得到的Q10与单独生长季或非生长季的数据推求出的Q10存在很大差异。Es和Q10的季节变异可能与树干体内非结构性碳水化合物的含量及形态的季节性变化存在一定关系,但还需要进一步验证[18]。另外,为了深入理解树干呼吸不同组分对温度变化的响应机理,学者们从生理上将树干呼吸划分为提供能量合成新细胞的生长呼吸和维持细胞生命活动的维持呼吸两个部分[19],并假设秋冬季树木停止生长后的树干呼吸即为维持呼吸,并认为维持呼吸对温度的响应在年际尺度上是恒定的[19]。以往大多数研究发现维持呼吸对温度的响应较为敏感,而生长呼吸则对温度并不敏感[20],故不同季节生长呼吸和维持呼吸所占的比例并不相同,也可能造成Q10和Es的季节性变异。此外,尽管Q10和Es作为生态系统过程模型的重要参数被广泛应用于预测未来气候变暖对陆地生态系统影响的模拟研究[21],但该参数的估算却仅依赖于树干组织内的氮素含量[3,8,15],而并未考虑Q10和Es的季节变异,从而导致对于陆地生态系统碳储量的估算存在很大的不确定性[4,6,18]。
毛白杨(Populustomentosa)作为华北平原区农田防护林的主栽树种之一,在该区域具有较大的分布面积和栽培规模。同时,农田防护林也是陆地生态系统的重要组成部分,但长期以来人们主要关注的是其经济价值和防护效益,而往往忽视其在陆地生态系统碳循环方面所起到的关键性作用。本研究以毛白杨为研究对象,连续监测毛白杨树干呼吸速率和树干温度变化,深入分析树干呼吸速率的昼夜变异以及季节性动态变化规律,旨在揭示影响毛白杨树干呼吸速率及其温度敏感性的潜在机理,以期为准确估算陆地生态系统碳储量和预测全球变暖进程提供基本的数据支撑和理论依据。
1 材料与方法
1.1 研究区概况
本研究样地位于河北省邯郸市南郊区的典型毛白杨农田防护林内(东经114°30′,北纬36°25′)。研究区域属典型暖温带半湿润大陆性季风气候,日照充足,雨热同期,四季的交替明显。该地区多年平均降雨量548.9 mm,主要集中在7—8月份,年平均气温14℃,最冷月份(1月)平均气温为-2.5℃,最热月份(7月)平均气温27℃,全年无霜期200 d,年日照2500 h。研究样地内的土壤质地以粉砂和轻壤为主;pH值较高,为7.5—8.0;土壤有机质含量较低,为6.0—10.0 g/kg; 全N含量为0.5—0.7 g/kg。
1.2 树干呼吸的测定
于2015年11月在试验区分别选取3个区域,在3个区域内分别选取生长良好的6棵毛白杨林木(共计18棵)用于树干呼吸速率和树干温度的连续监测(2016年1月份—2017年1月份,共计监测23次,具体日期见图3)。其中2016年4月26日(116 d)对树干呼吸进行了24 h的监测,其余的树干呼吸监测时间为8:00—20:00。
树干呼吸测定采用Li-Cor 6400便携式光合作用测量仪配套使用的呼吸测量气室。先将PVC环的一端切割成弧形以匹配树干的弧度,另一端磨平用以连接呼吸测量室。在选择的毛白杨树干1.5 m高度处安装PVC环(内径10 cm,深度5 cm)。安装前轻微刮掉表层树皮,尽量确保树干表面的平整性,但不能损伤到形成层组织。用硅胶将呼吸环固定在树干表面上,检查其密封性,确保不漏气。在呼吸环的右侧5 cm位置钻取一个细孔,使Li-Cor 6400自带的温度探头刚好插进树干,在监测树干呼吸的同时测量树干温度。
测定树干呼吸时,将Li-Cor 6400呼吸测量室扣在树干上固定的呼吸环,每间隔2—3 h对18棵毛白杨树干呼吸及树干温度进行1次测定。对18棵树干呼吸及树干温度取平均值,作为每次测定的均值;对每天内的数据取平均值作为当天测定的均值。样地内林木及其测定位置的基本特征见表1。根据毛白杨在本研究区的物候特征(展叶期为4月份,落叶期为11月份)将整个研究期划分为生长季(2016年5月份—10月份)和非生长季(2016年11月份—2017年1月份)。进入非生长季后树木开始落叶,研究区进入秋季,树木停止生长。本研究认为在非生长季测定的呼吸即是维持呼吸,而在生长季测定的呼吸则是生长呼吸和维持呼吸的总和[19]。

表1 毛白杨树干呼吸监测林木及样地特征
1.3 树干可溶性糖和淀粉的含量分析
每块样地附近随机选取2棵与测定树相似的样树,共计6棵白杨树,分别在生长季(2016年5月18日,第138天)和非生长季(2016年11月23日,第327天)使用生长锥钻取树芯样品,取样孔径12 mm,取样深度4 cm。考虑到树干内碳水化合物的日变化,故取样时间统一在9:00—11:00进行[1]。将树芯样品于烘箱内在75 ℃条件下烘干72 h至恒重。样品烘干后粉碎封闭储存。采用高压液相色谱法(HPLC)测定树芯样品中的淀粉和可溶性糖(蔗糖、葡萄糖、果糖)含量。
1.4 数据分析
为了计算树干呼吸速率,需要获得PVC环所围树干面积以及气室插入PVC环的有效深度。根据公式(1)计算PVC环所围的树干面积[18]。
(1)
A指PVC环所围的树干面积,Dc指PVC环的直径,Ds指树干直径。
根据公式(2)计算气室插入PVC环的有效深度[18]。
H=(Vc-(Dc/2)2πd)/(Dc/2)2π
(2)
H指有效插入深度,Vc和Dc指树干上PVC环的体积和直径。
本研究采用公式(3)拟合树干呼吸速率和树干温度之间的关系[22],按公式(4)计算温度敏感系数Q10[23]。
ES=β0eβ1T
(3)
式中:β0为当树干温度为零时的树干CO2释放通量;β1为温度系数。
Q10=e10β1
(4)
式中:Q10为树干温度每增加10℃呼吸速率增加的倍数。
为了消除温度的干扰,比较相同树干温度时树干呼吸速率在不同时间和季节上的差异,需将毛白杨树干呼吸统一矫正为15℃时的呼吸速率(R15)[21]。R15由树干呼吸和树干温度的回归方程(E-T曲线)得到。
本研究数据利用单因素方差分析的方法(ANOVA)比较林木组间的显著性差异(n=18)。所有的统计分析均利用SPSS 13.0 (Chicago, IL) 统计软件完成,所有作图均利用Excel软件实现。
2 结果与分析
2.1 毛白杨树干呼吸的日变化特征

图1 生长季树干温度和树干呼吸速率的日变化 (±标准误,n=18)Fig.1 Diurnal variation of stem respiration and stem temperature during growing season (±SE,n=18)
2016年4月26日(116天),对毛白杨树干呼吸及树干温度进行24 h监测,结果显示毛白杨树干温度呈现出先升高后降低的变化趋势,且树干温度的最高值为22.8℃,出现在下午13:00,最低值为15.3℃,出现在次日凌晨5:00时(图1)。然而,毛白杨树干呼吸速率的变化趋势同树干温度的变化并不一致,具体表现为树干呼吸速率的最大值出现在傍晚约19:00,树干呼吸速率最大值的出现相比树干温度最高值后滞了8 h,且树干呼吸速率在白天随时间推移呈现出微弱降低的趋势,从早晨7:00的1.0 μmol CO2m-2s-1逐渐降低为下午15:00的0.76 μmol CO2m-2s-1,尽管在下午17:00树干呼吸速率略微升高。本研究还发现毛白杨在夜间的树干呼吸速率明显高于白天,具体表现为毛白杨树干呼吸速率从下午17:00到傍晚19:00出现明显的跳跃,树干呼吸速率由大约0.8 μmol CO2m-2s-1突然升高到最高值1.42 μmol CO2m-2s-1,相比树干呼吸速率最低值增加87%(图1)。
对毛白杨树干温度和树干呼吸速率进行指数函数拟合的结果显示,毛白杨树干温度与树干呼吸速率存在较好的指数关系(E-T曲线),但在白天(7:00—19:00)和夜间(19:00—7:00)存在明显的差异(图2)。白天的毛白杨树干温度与树干呼吸速率呈现负相关的关系,树干温度解释了树干呼吸速率变化的88%。然而,在夜间树干呼吸速率与温度之间表现为正相关关系,树干温度解释了树干呼吸速率96%的变异(图2)。
2.2 毛白杨树干呼吸及温度敏感性的季节动态特征
毛白杨树干呼吸速率在季节尺度上同树干温度的变化趋势具有较好的一致性(图3)。在整个观测期内,毛白杨树干温度和树干呼吸速率均随季节变化呈现出先升高后降低的变化趋势,树干呼吸的变化范围为0.11—2.63 μmol CO2m-2s-1,树干温度的波动范围为1.8—33℃。毛白杨树干呼吸在春季(4月初)开始升高,并在夏季(8月末)达到最高值2.63 μmol CO2m-2s-1,而后随气温的降低逐渐下降,在冬季(12月中旬)达到最低值0.11 μmol CO2m-2s-1(图3)。在整个观测期内毛白杨树干呼吸速率同树干温度之间也存在着较好的指数函数关系(E-T曲线),树干温度可以解释毛白杨树干呼吸速率季节变异的93%,毛白杨树干呼吸的温度敏感性指数(Q10)为2.62(图4)。

图2 日变化尺度上树干呼吸和树干温度的关系Fig.2 Relationships between stem respiration and stem temperature during diurnal variation
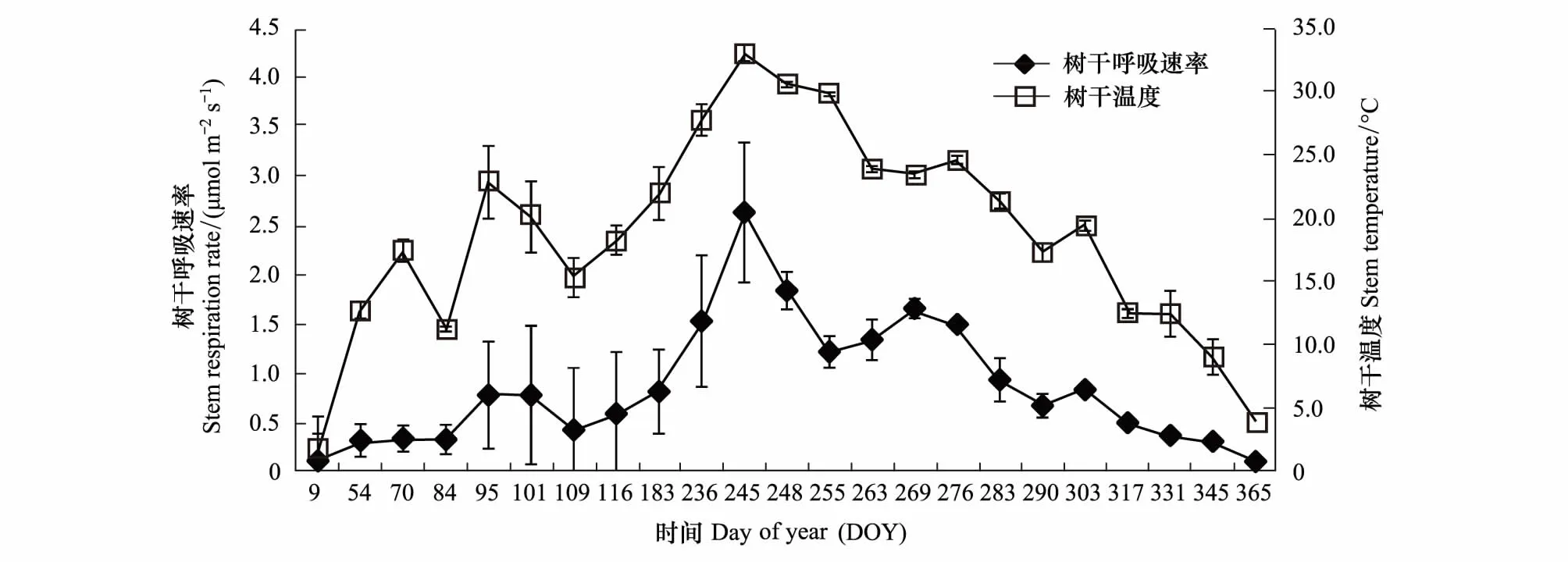
图3 树干呼吸和树干温度的季节变化(±标准误,n=18)Fig.3 Seasonal variation of stem respiration and stem temperature (±SE,n=18)

图4 整个研究期树干呼吸和树干温度的关系 Fig.4 Relationships between stem respiration and stem temperature during the observation periods
根据毛白杨物候特征,将整个研究期划分为生长季和非生长季,结果显示生长季和非生长季的树干呼吸和树干温度之间都存在指数函数关系(生长季R2=0.68,非生长季R2=0.79),表明树干温度在生长季和非生长季可以分别解释树干呼吸季节变化的68%和79%(图 5)。另外,本研究结果还显示,非生长季树干呼吸的温度敏感性系数(Q10=3.00)明显高于生长季(Q10=1.95)(图5)。
本研究结果显示,毛白杨树干呼吸在夜间的R15为0.58 μmol CO2m-2s-1,显著高于白天的R15值0.35 μmol CO2m-2s-1(P<0.01,图 6)。在季节尺度上R15的变化却与温度变化呈现比较一致的趋势。生长季毛白杨树干呼吸(生长呼吸与维持呼吸之和)的R15为0.65 μmol CO2m-2s-1,显著高于非生长季毛白杨树干呼吸(维持呼吸)的R15值0.52 μmolCO2m-2s-1(P<0.05,图 6)。
2.3 树干呼吸速率与树干非结构性碳水化合物的关系
本研究结果显示,毛白杨树干可溶性糖和淀粉的含量在不同季节间的差异显著,且生长季毛白杨树干可溶性糖及淀粉的含量均明显高于非生长季(P<0.05,图 7)。具体而言,生长季毛白杨树干的可溶性糖含量为86.69 mg/g,而非生长季的可溶性糖含量降低为63.83 mg/g,即毛白杨生长季的树干可溶性糖含量比非生长季增加36%。相似地,生长季毛白杨树干的淀粉含量为26.24 mg/g,明显高于非生长季的淀粉含量(18.24 mg/g)44%。
本研究分别拟合了生长季和非生长季毛白杨树干呼吸(R15)与树干可溶性糖或淀粉含量的线性关系(图 8)。拟合结果显示,就生长季而言,毛白杨树干的可溶性糖含量与R15的相关性(R2=0.52)高于淀粉(R2=0.35);相反,毛白杨树干的淀粉含量与R15在非生长季的线性关系(R2=0.59)相比可溶性糖含量与R15的相关性更好(R2=0.50)。
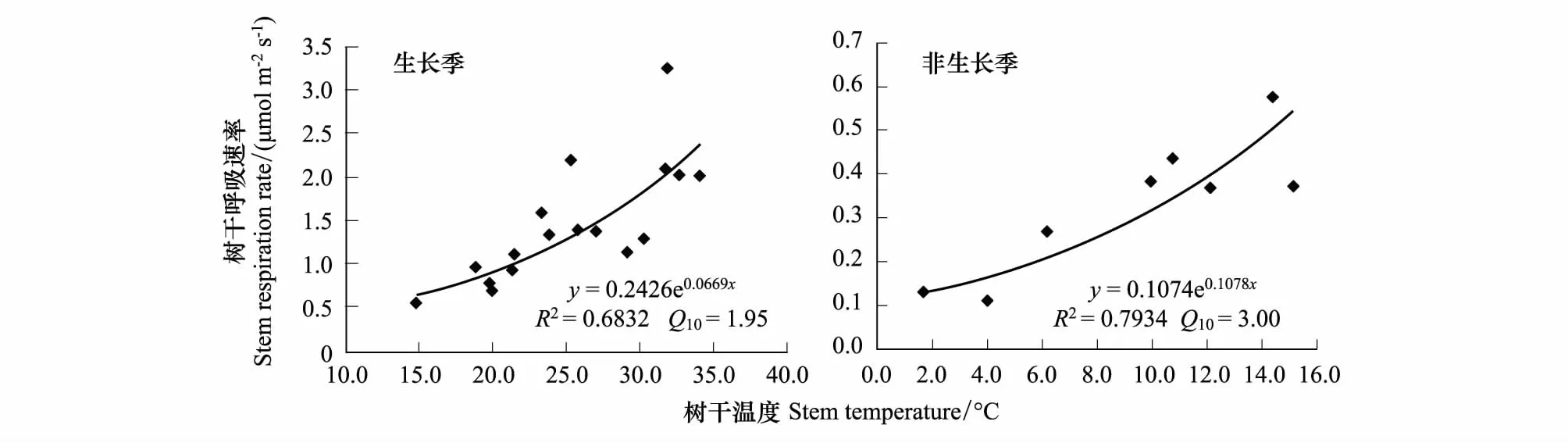
图5 生长季和非生长季树干呼吸与树干温度的关系Fig.5 Relationships between stem respiration and stem temperature during growing season and non-growing season

图6 温度矫正树干呼吸的昼夜变化和季节变异(±标准误,n=18)Fig.6 Diurnal and seasonal variations of temperature-normalised stem respiration (±SE, n=18)
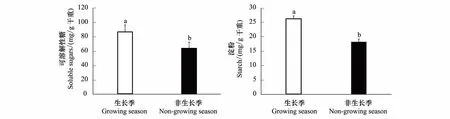
图7 树干可溶性糖和淀粉含量的季节变化(±标准误,n=6)Fig.7 Seasonal variations of the stem soluble sugar and starch contents (±SE,n=6)

图8 树干可溶性糖和淀粉浓度与R15在不同季节的关系Fig.8 Relationships of soluble sugars and starch with R15 in the growing season and non-growing season
3 讨论
3.1 树干呼吸日变化分析
树干呼吸日变化往往会受到树干温度和树干液流的影响,且树干温度和树干液流存在明显的昼夜变化[24],导致影响树干呼吸日变化的机理极其复杂,以至于目前对于森林碳储量的估算还存在很大的不确定性[8,10]。以往的大多数研究结果表明,森林树干呼吸日变化曲线为双峰型[19]和单峰形[25],且树干呼吸速率最大值出现在16:00—20:00时,而最小值则出现于4:00—10:00[25]。同时,另有相关研究结果发现树干呼吸通常滞后于树干温度约2—3个小时[26]。然而,本研究结果显示毛白杨树干呼吸昼夜动态变化呈单峰型曲线,其峰值出现在傍晚大约19:00时,且树干温度与树干呼吸在白天呈现出相反的变化趋势。以往的相关研究也发现类似的现象,Zhu等[27]在对木荷树干呼吸的研究中发现在7月份,树干呼吸和树干温度呈现出负相关的现象,并解释为由于高温抑制了树干呼吸作用,导致了树干呼吸速率的降低。王秀伟等[28]的研究也发现,3个树种(落叶松、水曲柳、白桦)的树干呼吸与树干温度在6月份呈现出相反的趋势,其原因可能是白天树干液流速度大,树干呼吸产生的部分CO2可能通过植物的蒸腾作用而进入大气中,故低估了树干呼吸的测量值,从而导致树干表面CO2释放量在夜间高于白天。本研究监测显示在4月份树干温度最大值为22.76℃,故不可能是高温抑制树干呼吸,而应该是白天树干液流带走部分CO2气体而造成的,即造成树干呼吸峰值出现在傍晚时的主要原因是由于白天树体内部生成的部分CO2气体被树干液流运输至叶片并通过蒸腾作用释放到大气中[29]。事实上,以往的大多数研究也发现树干呼吸与树干液流之间存在显著的负相关关系[27,30],树体内部细胞代谢过程产生的CO2气体并不会全部通过树皮扩散到外界大气中,而是一部分CO2气体溶解于树干液流中并向上运输,最终通过叶片气孔释放到大气环境,以至于许多研究发现树干呼吸峰值出现在温度较低的夜间[29,31]。另外,本研究分别拟合白天和夜间毛白杨树干呼吸对温度响应的曲线,结果显示树干呼吸和树干温度在夜间的相关系数高于白天,这可能是由于夜间受到液流的干扰小,树干呼吸对温度的响应更加敏感,而白天液流对树干呼吸的干扰较大,从而降低了树干呼吸对温度响应的敏感性。然而,液流对树干呼吸速率的影响程度在不同季节、不同气候条件甚至不同树种间存在很大的变异[27,29]。因此,目前关于液流对树干呼吸产生影响的潜在机理还存在较大争议[27,28],有待于在不同森林类型中更进一步的深入探讨和研究。
3.2 树干呼吸及温度敏感性的季节变化规律
在整个观测期内树干呼吸呈现出明显的季节变化规律(图3),且同树干温度具有很好的相关性(R2=0.93,图4),表明在季节变化的尺度上树干温度可能是控制树干呼吸的主要因素之一,因为在较高树干温度时树体内部细胞的线粒体呼吸速率增大,从而增加CO2气体的释放量,而且还会提高CO2气体从树体内部到外界大气的扩散系数[21]。另外,本研究分别拟合生长季和非生长季E-T曲线的结果表明,相对于生长季而言,非生长季毛白杨树干呼吸与温度之间具有更好的指数回归关系(非生长季时的R2高于生长季),该结果与Ryan的研究结论一致[32]。本研究结果表明,在非生长季树干呼吸全部为维持呼吸,维持呼吸对温度的响应更加敏感,故其相关性更高,回归系数更大。然而,在生长季所测树干呼吸为生长呼吸与维持呼吸之和,生长呼吸对温度的响应不敏感。因此,生长季树干呼吸与温度的回归系数较小。同样,王淼等的研究也发现幼树的树干呼吸与温度的相关性比老树较差,主要是由于幼树处于快速生长阶段,故其生长呼吸部分占总呼吸的比例较大,从而导致树干呼吸与树干温度相关性的下降[25]。此外,以往大多数研究结果表明树干呼吸温度敏感性(Q10)随着物种、气候条件、底物供应状况等的变化而发生改变[30,33],树干呼吸Q10值的变化范围为1.0—4.4[34-35]。本研究结果显示,毛白杨树干呼吸非生长季的Q10值(3.00)高于生长季的Q10值(1.95),非生长季树干呼吸主要为维持呼吸,这也表明维持呼吸对温度的响应更加敏感。然而,利用整个观测期数据拟合得到的Q10为2.62,与在生长季和非生长季拟合出的Q10也存在很大的不同,表明在不同的时间尺度上,利用E-T曲线拟合出的温度敏感指数存在很大的变异。因此,基于生态系统过程模型预测森林生态系统碳循环过程对气候变暖响应的相关研究应该充分考虑到森林树干呼吸过程在不同季节(生长季和非生长季)响应温度变化存在差异的情况。
3.3 可溶性糖与淀粉的季节变化对树干呼吸的影响
毛白杨生长季的R15显著高于非生长季,表明除温度以外的其他因素也可能在毛白杨树干呼吸季节变化中起作用。本研究对不同季节树体内非结构性碳水化合物含量(可溶性糖和淀粉)测定的结果显示,生长季毛白杨树干中的可溶性糖和淀粉含量均显著高于非生长季,表明毛白杨生长季R15高于非生长季的原因可能是由于生长季可利用的呼吸底物(非结构性碳水化合物)含量比非生长季更高而造成的。以往的一些研究结果也表明非结构性碳水化合物含量与树干呼吸之间存在着密切的相关性[21,36]。例如,Maier等发现火炬松树体组织中的非结构性碳水化合物含量与树干呼吸存在明显的正相关关系[21]。在生长季树干中可溶性糖与R15存在更好的相关关系;而在非生长季树干中的淀粉与R15存在更好的相关关系。这表明在非生长季树干呼吸的主要底物形式为淀粉,树体内储存的大量淀粉物质被消耗用于林木维持呼吸过程,以至于树干组织的非结构性碳水化合物含量降低[37];然而,生长季树干呼吸的底物主要以可溶性糖为主,且林木通过光合作用过程生成大量非结构性碳水化合物[37],并迅速以可溶性糖的形式运送至树体用于树干呼吸过程,进一步为林木新陈代谢过程提供能量。
4 结论
本研究发现毛白杨树干呼吸的Q10在不同季节存在明显变异,且生长季Q10值(1.95)显著低于非生长季Q10值(3.00),表明毛白杨树干呼吸不同组分对温度响应的敏感程度并不相同。另外,生长季树体内的可溶性糖含量与树干呼吸具有较好的相关性,但非生长季淀粉含量同树干呼吸的相关性更加明显,表明呼吸底物类型及其供应状况(非结构性碳水化合物)在一定程度上决定着毛白杨的树干呼吸速率及其温度敏感性,即毛白杨树干呼吸的温度敏感性(Q10)其实并非为呼吸过程对温度的真实响应,而是受其他多种因素(如呼吸底物类型、底物供应状况等)和生理过程共同影响的表观温度敏感性(ApparentQ10)。因此,利用生态系统过程模型估算森林生态系统呼吸非常有必要将树干总呼吸区分为生长呼吸和维持呼吸,并分别计算两者的温度敏感性,避免利用树干总呼吸的表观Q10直接估算森林生态系统呼吸而带来的误差,以至于高估气候变暖对树干呼吸过程的影响,进而低估森林生态系统呼吸对未来气候变暖的反馈能力。
猜你喜欢
军事文摘(2021年16期)2021-11-05
新农业(2021年15期)2021-08-23
新农民(2020年21期)2020-12-08
河北林业科技(2020年3期)2020-01-10
东坡赤壁诗词(2018年1期)2018-03-31
小猕猴学习画刊(2017年1期)2017-02-17
小猕猴学习画刊(2017年1期)2017-02-17
中国质量与标准导报(2014年5期)2014-02-28
储能科学与技术(2014年5期)2014-02-27
储能科学与技术(2014年1期)2014-02-27
