
人工老化对沙葱种子中活性氧与超微弱发光的影响
2019-11-07 11:15
种子 2019年10期
(内蒙古农业大学园艺与植保学院,内蒙古野生特有蔬菜种质资源与种质创新重点实验室, 呼和浩特 010019)
种子老化(Seed aging)是指种子生存能力下降,使得种子丧失活力及萌发力的不可逆转变化,是在种子成熟后发生和发展的、自然而不可避免的过程,其起因与有害物质积累、内源激素紊乱、生物大分子变性和膜系统损伤等有关。目前,关于种子老化的具体机制尚不清楚,一般认为老化过程会表现出种子活力降低,膜脂过氧化,保护酶、蛋白质和核酸降解等特点[1]。因此,探究种子老化机理及如何缓解种子老化意义重大。研究发现,种子在生产中肥料供给充足,种子贮藏时降低种子含水量,低温、低湿贮藏等方法可以有效缓解种子老化[2]。沙葱种子贮藏过程中易发生老化劣变,在老化过程中活性氧和超微弱发光如何响应,是否可通过超微弱发光的强弱来无损伤的鉴定种子活力以及如何缓解种子的老化等问题未见报道。因此,本试验通过铈离子+人工老化处理沙葱种子,研究铈离子对沙葱老化种子的活性氧与超微弱发光的影响,以期为沙葱种质资源的保存提供科学依据。
1 材料与方法
1.1 试 材
以2016年采自鄂尔多斯市鄂托克前旗荒漠草甸的沙葱(AlliummongolicumRegel.)种子为试材,千粒重为(2.28±0.06)g,初始含水量为(5.78±0.33)%。
1.2 浸种处理
选择饱满、大小一致、无破损的沙葱种子,经400 mg·L-1Ce3+(氯化亚铈)浸种6 h后铺在滤纸上室温晾干。
1.3 人工老化处理
采用高温高湿法对沙葱种子进行人工老化[3],设定老化箱温度50 ℃,相对湿度(RH)为100%。老化处理时间分别为0,5,15,25 h和35 h。经Ce3+浸种后再人工老化处理分别记为Ce3+0、Ce3+1、Ce3+2、Ce3+3、Ce3+4;未经Ce3+浸种直接老化的沙葱种子分别记为ck0、ck1、ck2、ck3、ck4;老化完毕后取出种子,在室温下晾干至其原始水分,然后密封保存在4 ℃冰箱中备用。
1.4 萌发指标测定
取处理好的沙葱种子,用1%的次氯酸钠溶液杀菌消毒10 min,蒸馏水冲洗干净后放在铺有2层滤纸的培养皿(d=9 cm)中进行萌发试验,恒温培养箱温度为(20±1)℃,每皿50粒种子,4次重复。每隔24 h记录发芽数并补水,第10天结束试验,统计发芽率和发芽势,测定鲜重以及胚根长,计算发芽指数和活力指数。
发芽势(%)=(第3天发芽种子数/测定种子总数)×100%[4];
发芽率(%)=(发芽种子数/测定种子总数)×100%[4];
发芽指数=∑Gt/Dt,Gt为t时间内的发芽数,Dt为相应的发芽日数[4];
活力指数=发芽指数×鲜重。
1.5 生理指标测定

超微弱发光的测定使用超微弱发光测试系统(BPCL-2-SH),共测5次,减去本底取平均值,即为沙葱种子的最终发光强度。
1.6 数据分析
用 Microsoft Excel 2007 软件作图,差异显著性用Duncan’s检验,SPSS 17.0 软件进行相关性分析。
2 结果与分析
2.1 人工老化对沙葱种子萌发及活力的影响
由表1可知,沙葱种子各萌发指标随着老化时间的延长呈降低的趋势。ck0的各发芽指标显著高于ck1、ck2、ck3、ck4,在老化35 h后,活力指数为1%,表明沙葱种子几乎失活。而Ce3+处理下,Ce3+1各发芽指标值高于ck1,平均发芽率比ck1高4%,活力指数比ck1高0.042,平均胚根长比ck1长1.64 cm。表明Ce3+有助于提高老化初期沙葱种子活力及促进胚根的伸长。
2.2 人工老化对沙葱种子相对电导率的影响
由图1可知,沙葱种子相对电导率随着老化时间的延长呈先增大后减小再增大的趋势,但各处理间差异不显著,其中ck1相对电导率高于其他处理,表明种子随老化胁迫种子结构发生改变,内部物质外渗。而Ce3+处理,电导率随着老化时间的延长呈先减小再增大的趋势;在老化处理5 h之前,Ce3+处理后的电导率值低于ck,随后老化胁迫作用大于Ce3+处理作用,种子相对电导率随对照逐渐升高。表明在老化程度较低的情况下,Ce3+前处理对沙葱种子电解质外渗有一定缓解作用。

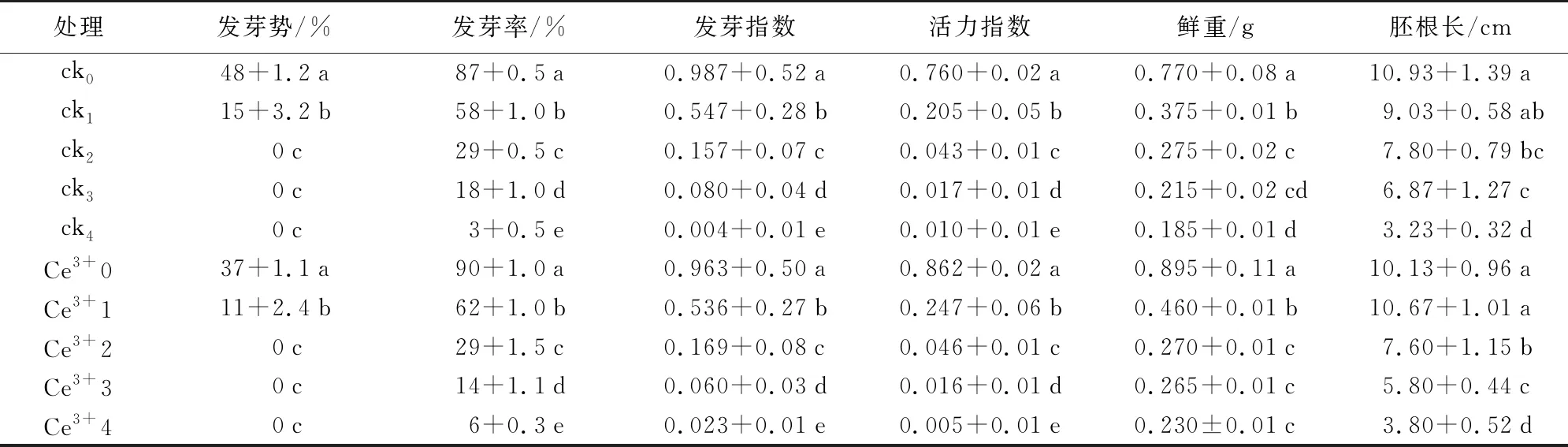
处理发芽势/%发芽率/%发芽指数活力指数鲜重/g胚根长/cmck048+1.2a87+0.5a0.987+0.52a0.760+0.02a0.770+0.08a10.93+1.39ack115+3.2b58+1.0b0.547+0.28b0.205+0.05b0.375+0.01b9.03+0.58abck20c29+0.5c0.157+0.07c0.043+0.01c0.275+0.02c7.80+0.79bcck30c18+1.0d0.080+0.04d0.017+0.01d0.215+0.02cd6.87+1.27cck40c3+0.5e0.004+0.01e0.010+0.01e0.185+0.01d3.23+0.32dCe3+037+1.1a90+1.0a0.963+0.50a0.862+0.02a0.895+0.11a10.13+0.96aCe3+111+2.4b62+1.0b0.536+0.27b0.247+0.06b0.460+0.01b10.67+1.01aCe3+20c29+1.5c0.169+0.08c0.046+0.01c0.270+0.01c7.60+1.15bCe3+30c14+1.1d0.060+0.03d0.016+0.01d0.265+0.01c5.80+0.44cCe3+40c6+0.3e0.023+0.01e0.005+0.01e0.230±0.01c3.80+0.52d

图1 人工老化过程中沙葱种子相对电导率变化
2.3 人工老化对沙葱种子呼吸速率的影响
由图2可知,沙葱种子呼吸速率随着老化时间的延长呈先增大后减小的趋势,ck1呼吸速率最大,为0.12μmol·(g·min)-1,显著高于其他处理。Ce3+处理组与ck有相似趋势,Ce3+1处理呼吸速率最大。在老化15 h之前,Ce3+处理结果低于ck,在老化处理15 h之后,Ce3+处理结果又高于ck。在老化5 h到15 h时,呼吸速率降低的最快,表明在此阶段物质消耗最多。

图2 人工老化过程中沙葱种子呼吸速率变化
2.4 人工老化对沙葱种子活性氧的影响
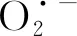

图3 人工老化过程中沙葱种子超氧阴离子产生速率的变化
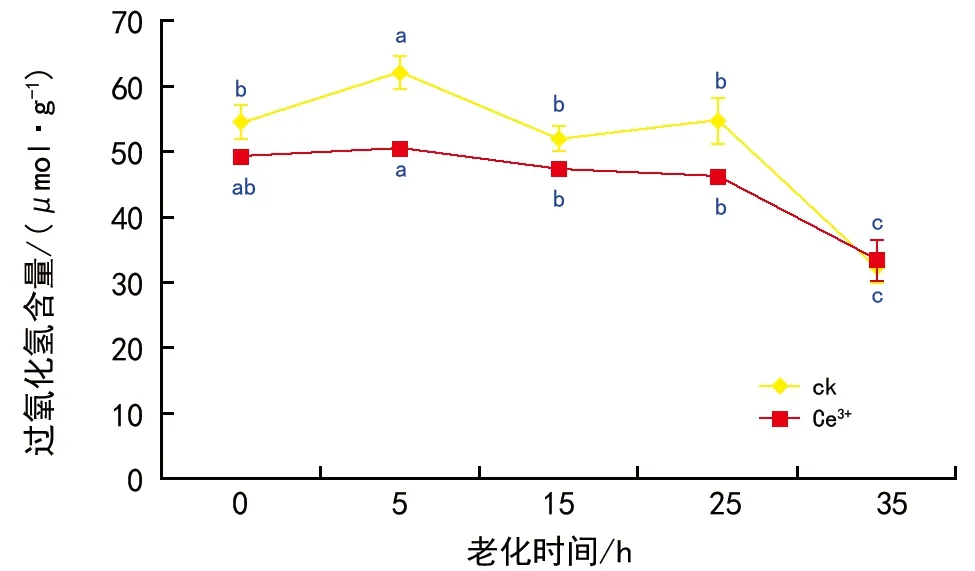
图4 人工老化过程中沙葱种子过氧化氢含量的变化
2.5 人工老化对沙葱种子超微弱发光的影响
由图5可知,沙葱种子超微弱发光强度随老化时间的延长呈逐渐降低的趋势,各处理间差异显著,ck2、ck3和ck4比ck0分别降低了30%、37.3%和73.3%。
表2 沙葱种子活力与部分生理指标间的相关性
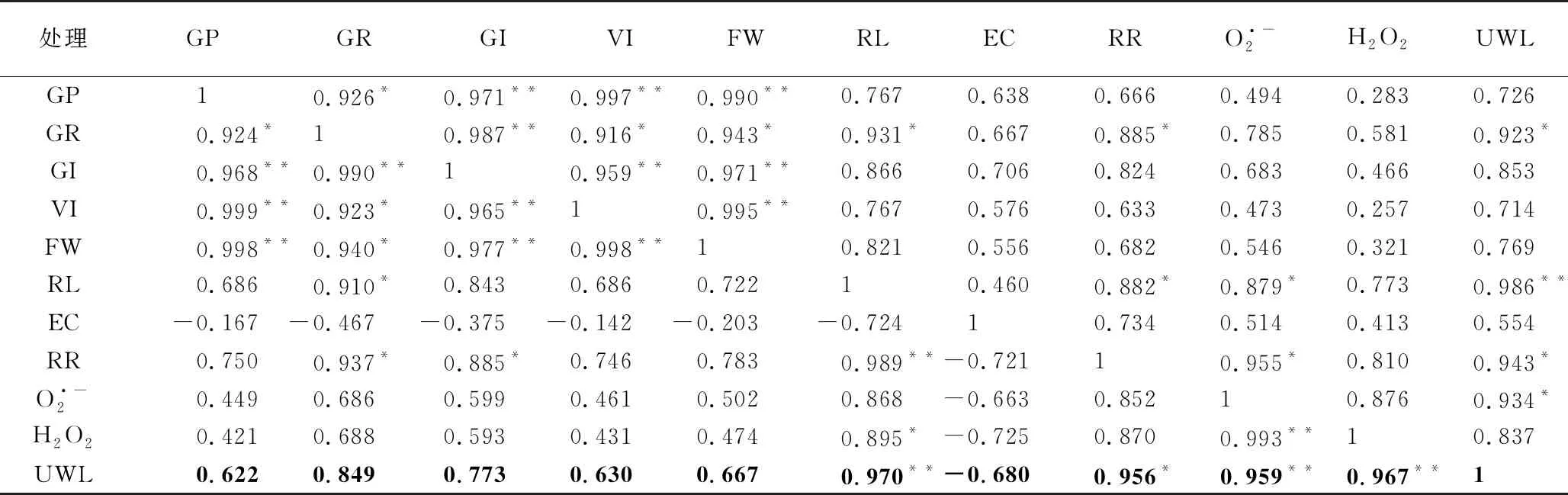
处理GPGRGIVIFWRLECRRO·-2H2O2UWLGP10.926*0.971**0.997**0.990**0.7670.6380.6660.4940.2830.726GR0.924*10.987**0.916*0.943*0.931*0.6670.885*0.7850.5810.923*GI0.968**0.990**10.959**0.971**0.8660.7060.8240.6830.4660.853VI0.999**0.923*0.965**10.995**0.7670.5760.6330.4730.2570.714FW0.998**0.940*0.977**0.998**10.8210.5560.6820.5460.3210.769RL0.6860.910*0.8430.6860.72210.4600.882*0.879*0.7730.986**EC-0.167-0.467-0.375-0.142-0.203-0.72410.7340.5140.4130.554RR0.7500.937*0.885*0.7460.7830.989**-0.72110.955*0.8100.943*O·-20.4490.6860.5990.4610.5020.868-0.6630.85210.8760.934*H2O20.4210.6880.5930.4310.4740.895*-0.7250.8700.993**10.837UWL0.6220.8490.7730.6300.6670.970**-0.6800.956*0.959**0.967**1
注:GP为发芽势;GR为发芽率; GI为发芽指数;VI为活力指数;FW为鲜重;RL为胚根长;EC为电导率;RR为呼吸速率。*表示在0.05水平显著相关;**表示在0.01水平极显著相关。
在Ce3+处理组中,Ce3+比ck的超微弱发光强度高,Ce3+1和Ce3+0的超微弱发光强度差异不显著,Ce3+0显著高于Ce3+2、Ce3+3和Ce3+4的超微弱发光强度,Ce3+2、Ce3+3和Ce3+4比Ce3+0分别降低了18.4%、32%和68%。

图5 人工老化过程中沙葱种子超微弱发光的变化
2.6 相关性分析

3 讨 论
3.1 人工老化对沙葱种子活性氧的影响
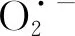
3.2 人工老化对沙葱种子超微弱发光的影响
超微弱发光( Ultraweak luminescence,UWL)检测技术可以在不破坏生物体时探究生物体生理信息,研究发现超微弱发光与生物机体的氧化代谢、细胞分裂和死亡、细胞内和细胞间的信息传递以及生长过程等有着密切的联系[19-20]。研究发现,小麦种子随着老化程度的加深,超微弱发光强度逐渐降低[21-22]。在ck处理组,超微弱发光与发芽率呈显著正相关。这与张文兰等[23]的研究结果一致。而庞靖祥等研究发现,板蓝根种子的发芽率与其自发发光的强度是负相关的[24]。本试验中发现,在ck 处理组,超微弱发光强度与相对电导率呈正相关,在Ce3+处理组,两者呈负相关。这与田茜等[25]的研究结果一致,表明Ce3+有效作用细胞的生物膜系统,有效的缓解了细胞膜的破坏和种子内含物外渗。

图6 苹果酸调控植物程序性细胞死亡模式图
综上所述,在种子的加速老化过程中,超微弱发光强度与活性氧、呼吸作用、发芽指标等生理指标有一定相关性,Ce3+处理可调控活性氧的增加,减缓老化对细胞损伤,维持各项功能稳定性。超微弱发光强度与活性氧呈极显著相关,表明超微弱发光强度随活性氧的变化而变化,与生物化学提出的“活性氧”机制相一致。然而,Ce3+是如何调控活性氧迸发,缓解沙葱种子老化,以及在生物变化的某个过程或影响生物某个代谢循环需要进行深入的分析;另外,本试验发现在老化5 h是Ce3+处理和ck在相对电导率高低的分界点,细胞受胁迫的程度可通过测定细胞电解质渗出率反应, 电导率数值的大小反应细胞膜透性的变化, 两者的变化趋势一[26],因此将在接下来的细胞膜透性试验中验证。相关系数分析得出超微弱发光与超氧阴离子产生速率和过氧化氢含量呈极显著正相关,研究发现线粒体组分参与PCD表明线粒体也能触发和执行PCD过程,ROS可以充当诱导PCD的信号分子,改变线粒体膜通透性,细胞色素C和线粒体电子传递链(mETC)崩解释放的ROS足以促使PCD的发生达到致死水平[27]。可假想种子老化过程ROS的产生可诱导种子PCD(图6),又UWL相关ROS,推测在细胞外UWL反映PCD形态特征,在细胞内ROS反映PCD生理特征。在拟南芥中, 叶绿体中生成的苹果酸通过DiT 1运输至细胞质,继而被某个/某些转运蛋白转运至线粒体。苹果酸在线粒体中被mMDH 1催化形成草酰乙酸,并伴随着NADH的积累,为线粒体膜上的mETC复合体提供电子,产生ROS,进而导致产生程序性细胞死亡[28]。
猜你喜欢
现代农业科技(2022年5期)2022-12-14
石油库与加油站(2022年3期)2022-08-30
科学导报(2022年11期)2022-03-03
矿冶工程(2021年4期)2021-09-15
智慧健康(2020年9期)2020-12-03
世界有色金属(2020年15期)2020-10-10
上海农业科技(2020年2期)2020-04-02
山西农业科学(2020年1期)2020-02-27
农民致富之友(2018年9期)2018-06-27
湖北农业科学(2014年13期)2014-08-28
