
不同供钾水平下烤烟生长及硝态氮吸收动力学特征
2019-10-22 01:18林昌华张士荣肖洲刘春玲李璐瑶王军丁效东
热带作物学报 2019年9期
林昌华 张士荣 肖洲 刘春玲 李璐瑶 王军 丁效东
摘 要 采用盆栽砂培試验方法研究了不同钾水平供应下烤烟生长及其对吸收硝态氮的动力学参数特征的影响。主要结论如下:与低钾(K1)、高钾(K3)处理相比,烤烟在中钾(K2)处理时地上部、根系鲜、干重均增加;随钾浓度增加,钾由根系向地上部运输比例增加,而烤烟生育前期净光合速率无显著性差异,但后期净光合速率显著增加;中钾(K2)水平增加根系总根长、总表面积、平均直径和总体积,而高钾(K3)水平反而有抑制作用。不同供钾水平预处理后,烤烟对NO3--N吸收符合离子吸收动力学模型,其吸收动力学参数表现为钾水平预处理浓度增加后,Vmax增大,Km值增加,但增加的幅度不一致。与低钾(K1)预处理相比,高钾(K3)预处理降低烤烟根系高亲和力系统对NO3--N的吸收速率,而中钾(K2)处理提高烤烟根系高亲和力系统的NO3--N的吸收速率,表明烤烟根系吸收NO3--N的高亲和力系统受钾水平调控,适宜钾浓度预处理提高烤烟根系对低浓度NO3--N的吸收。对低亲和系统来说,与低钾预处理相比,中钾(K2)和高钾(K3)预处理均提高了烤烟根系低亲和力系统对NO3--N吸收,表明长期高钾环境生长下的烤烟,根系对硝态氮吸收的低亲和力系统影响较小,而长期低钾环境生长下的烤烟,根系对硝态氮吸收低亲和力系统有抑制作用。
关键词 烤烟;钾预处理;硝态氮吸收;动力学特征;亲和力系统中图分类号 Q949.748.5 文献标识码 A
The Absorption Dynamic Parameter of NO3- in Flue-cured Tobacco Pretreated in Different Concentrations of KCl
LIN Changhua1, ZHANG Shirong2, XIAO Zhou2, LIU Chunling2, LI Luyao2, WANG Jun3, DING Xiaodong2*
1. Henry Fok School of Agricultural Science and Engineering, Shaoguan University, Shaoguan, Guangdong 512005, China; 2. College of Resources and Environment, Qingdao Agricultural University, Qingdao, Shandong 266109, China; 3. Nanxiong Tobacco Science Institute, Nanxiong, Guangdong 512400, China
Abstract In the present study, the growth and the absorption dynamic parameter of NO3--N in flue-cured tobacco, which was pretreated in different concentrations of KCl, were investigated with sand culture test. The results showed that compared with low (K1) and high K (K3) treatments, the fresh and dry weight of the shoot and root of the flue-cured tobacco increased under medium potassium (K2) treatments. With the increase of K concentration, the proportion of potassium transported from roots to shoots increased, while the net photosynthetic rate of the flue-cured tobacco had no significant difference in the early growth stage, but increased significantly in the late growth stage. Compared with low (K1), medium K (K2) level increased total root length, total surface area, average diameter and total volume, while high potassium (K3) level inhibited it; the uptake of NO3--N of the flue-cured tobacco was in accordance with Michelis-Menten equation pretreated with different concentrations of KCl, and the uptake kinetics parameters,Vmax andKm were enhanced with the K pretreatment concentration increased, but the increase range was not consistent. For the high-affinity system, compared with low K (K1) pretreatment, high K (K3) pretreatment decreased the uptake rate of NO3--N, while medium K (K2) pretreatment increased the uptake rate of NO3--N, which indicated that the high affinity system of NO3--N uptake by the flue-cured tobacco was regulated by K level, and appropriate K level pretreatment increased the uptake rate of low concentration NO3--N. For the low-affinity system, compared with low K pretreatment, medium K (K2) and high K (K3) pretreatment could significantly increase the absorption of NO3--N, which indicated that the root uptake of NO3--N of the low-affinity system was less affected by long-term high K environment, while was restrained at long period of low K environment.
Keywords flue-cured tobacco; K Pretreated; NO3--N uptake; kinetics characteristic; affinity system
DOI10.3969/j.issn.1000-2561.2019.09.002
煙草(Nicotiana tabacumL.)是需钾量较高的叶用经济作物,生产100 kg烟叶所需要的氮、磷、钾养分量为:氮(N)3.5 kg、磷(P2O5)0.6 kg、钾(K2O)7.5 kg[1]。随着烤烟集约化生产水平不断提高,钾成为烤烟作物产质量提高的限制因子,充足的钾素供应是获得优质适产烟叶的重要条件[1-2]。国际上常以较高的烟叶含钾量作为评定优质烟叶的重要指标,然而我国生产的烟叶钾素含量平均小于2%,低于国际优质烟叶2.5%的钾含量标准,烟叶含钾量的提高成为改善我国烤烟质量的关键措施[2]。缺钾后植物20%的生理活动及代谢过程发生变化相关,其中37%的代谢过程为氮代谢过程,钾与氮同化过程关系极为密切[3-4]。缺钾后作物体内中NO3--N减少,游离氨基酸增加,高N/C比氨基酸和中性氨基酸如谷氨酰胺、天冬酰胺、精氨酸、组氨酸、赖氨酸显著增加[5]。钾离子和NO3--N在植物体内运输时具有协同作用,钾转运蛋白和NO3--N转运蛋白在转录组水平上被外界环境共同调控,钾可影响到植物NO3--N的吸收和转运[6-7],缺钾后根系和地上部NO3--N含量出现一定程度的降低[8-9]。
硝酸盐(NO3-)是作物生长主要氮源,根系对其吸收是主动吸收过程,NO3--N进入细胞是逆电化学势梯度进行的,是依靠代谢能量ATP的吸收过程[6]。由细胞膜硝酸盐转运体,即高亲和力转运系统(系统Ⅰ)和低亲和力转运系统(系统Ⅱ)完成[6]。系统Ⅰ符合酶吸收动力学特性,其底物NO3--N饱和值为0.2~0.5 mmol/L,Km值介于10~100 μmol/L之间。作物根系内不同的NO3--N转运系统,在底物较宽范围下(5 μmol/L~50 mmol/L)能够吸收介质中的NO3--N[6]。NO3--N吸收的生理学研究已经证明植物体有一系列不同的运输载体存在, 其生理特性的差异主要表现在动力学、定位、可诱导性及作用机理方面[10]。吸收动力学参数(Km,Vmax)可在一定程度上衡量植物根系对离子的吸收能力[11]。前人针对不同供钾条件下烤烟根系生长进行了研究,而针对钾供应水平对烤烟根系对硝态氮吸收过程及动力学特征鲜见报道[12]。本研究通过不同供钾水平对烤烟根系生长的预处理,研究了烤烟根系对硝态氮的吸收能力,明确其吸收动力学参数特征,以期为理解钾供应水平(胁迫)烤烟吸收氮素能力及施肥提供依据。
1 材料与方法
1.1材料
供试烤烟为京烟48,由中国农业科学院烟草研究所提供。
1.2方法
1.2.1 试验设计 试验安排在青岛农业大学植物营养温室,采用盆栽砂培试验。于2017年3月进行漂浮育苗,于2017年5月进行移栽,移栽到盛有石英砂的塑料盆,每盆3株,塑料盆规格为上沿直径25 cm,高25 cm,石英砂总重6 kg。移栽后用营养液进行浇灌,营养液采用单子叶营养液,基本营养液配方(mol/L):Ca(NO3)2×4H2O 2.0×10-3,MgSO4×7H2O 0.65×10-3,H3BO31.0×10-6,MnSO4×H2O 1.0×10-6,ZnSO4×7H2O 1.0×10-6,CuSO4×5H2O 1.0×10-7,(NH4)6Mo7O24×4H2O 5.0× 10-9,EDTA-NaFe 0.1×10-3。
砂培试验初始营养液为1/2浓度的上述营养液,三叶一心后为完全营养液。之后在钾浓度梯度为:0.01 mmol/L(低钾,K1),12.5 mmol/L(中钾,K2),25.0 mmol/L(高钾,K3)的水平下连续28 d进行不同钾浓度梯度水平营养液浇灌处理。将处理28 d后的烤烟幼苗进行氮素吸收动力学参数测定,即把经饥饿处理(仅向烟苗浇灌蒸馏水)2 d的植株分别移入含有200 mL溶液的三角瓶中(每瓶1株),每个硝态氮吸收浓度重复3次,吸收2 h之后取出,冲洗根系,分别称根系和地上部鲜重,采用流动分析仪测定吸收前后溶液中NO3--N浓度的变化,根据吸收前后溶液NO3--N浓度的变化量计算单位鲜根重在单位时间内的N净吸收量,即根系对N的净吸收速率。
1.2.2 地上部和根系鲜重、干重测定 将现取植株的地上部和根系分开,离子水冲洗,吸干,称重,记录鲜重;然后将植物鲜样放在烘箱中105 ℃杀青30 min,80 ℃烘72 h,称重,记录干重。
1.2.3 净光合速率和胞间CO2浓度的测定 利用汉莎科学仪器有限公司CIRAS-3超便携式光合仪分别在烤烟幼苗培养至14、28 d时进行净光合速率[CO2μmol/(s·m2)]和胞间CO2浓度的原位测定。
1.2.4 根系总根长、表面积、总体积和平均直径测定 取收获后的烤烟根系,利用根系扫描仪(LA1600+scanner,Canada)扫描根系形态,用Win RHIZO根系分析系统分析根系数(根系总根长、表面积、总体积和平均直径)。
1.2.5 烤烟全钾含量测定 采用H2SO4-H2O2消化—火焰光度计法测定。
1.2.6 硝态氮吸收动力学参数测定 硝态氮的吸收速率参数测定采用(NO3--N)浓度梯度法。Ca(NO3)2浓度分别为0.05、0.1、0.2、0.4、0.6、1.0、2.0、4.0、6.0、8.0、10.0 mmol/L。把经氮饥饿处理2 d的植株洗净移入100 mL烧杯中(每杯1株),每个氮吸收浓度重复3次,吸收2 h之后取出,冲洗根系,分别称根系和地上部鲜重,采用全自动流动分析仪测定硝态氮含量,根据吸收前后溶液NO3--N浓度的变化量计算单位鲜根重在单位时间内的N净吸收量,即根系对N的净吸收速率。采用Michaelis-Menten方程的Hofstee转换式处理数据,求出吸收动力学参数Vmax(最大吸收速率)和Km(表观米氏常数)。
1.3数据处理
1.3.1 硝态氮吸收速率的计算 烤烟氮吸收速率即为单位烤烟根系鲜重吸收氮的速率,用下式计算:V=(C0–Ct)·U/(t·m),式中:V为氮的吸收速率,μmol/(g·h);C0为试验结束时培养液中的氮含量,μmol/L;Ct为试验结束时培养液中的氮含量,μmol/L;U为营养液体积,L;t为试验时间,h;m为烤烟根系鲜重,g。
1.3.2 數据处理 采用SPSS软件和Office 2010软件进行数据处理。
2 结果与分析
2.1不同施钾量对烤烟根系和地上部生长的影响
由表1可知,与低钾(K1)和高钾(K3)相比,烤烟幼苗在中钾(K2)处理时地上部鲜重显著增加,分别增加37.7%、34.0%;与低钾(K1)和高钾(K3)相比,烤烟幼苗在中钾(K2)处理时地上部干重显著增加,分别增加33.4%、62.1%。根系鲜干重与地上部表现一致,干重分别增加53.8%、55.5%;鲜重分别增加31.9%、38.8%。结果表明在不同钾水平梯度中,中钾(K2)水平处理的烤烟地上部鲜干重及根系鲜干重均有显著提升,即适当增加钾的施用量有利于增加烤烟地上部和根系的生物量,提高烤烟产量。
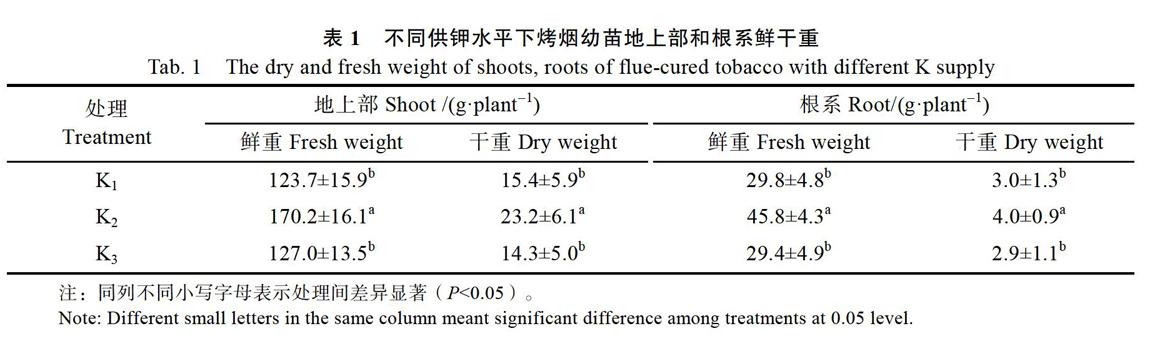
2.2不同施钾量对烤烟幼苗地上部和根系钾含量的影响
与低钾(K1)处理相比,烤烟幼苗在中钾(K2)处理及高钾(K3)处理时地上部钾含量显著增加,而中钾(K2)处理和高钾(K3)处理时地上部钾含量差异不显著(图1A);根系钾含量与地上部表现相同趋势(图1B)。结果表明,施钾能够增加烤烟地上部及根系钾含量,但施钾量增加到一定程度时地上部和根系钾含量变化不显著,即在一定钾浓度范围内,烟叶全钾含量与施钾用量存在显著正相关;随钾浓度增加,根系与地上部钾含量比值分别为3.54、3.48、2.97,表明随钾浓度增加,烤烟钾素由根系向地上部运输比例增加,即烤烟在高钾水平下,其烤烟地上部钾吸收速率提高。
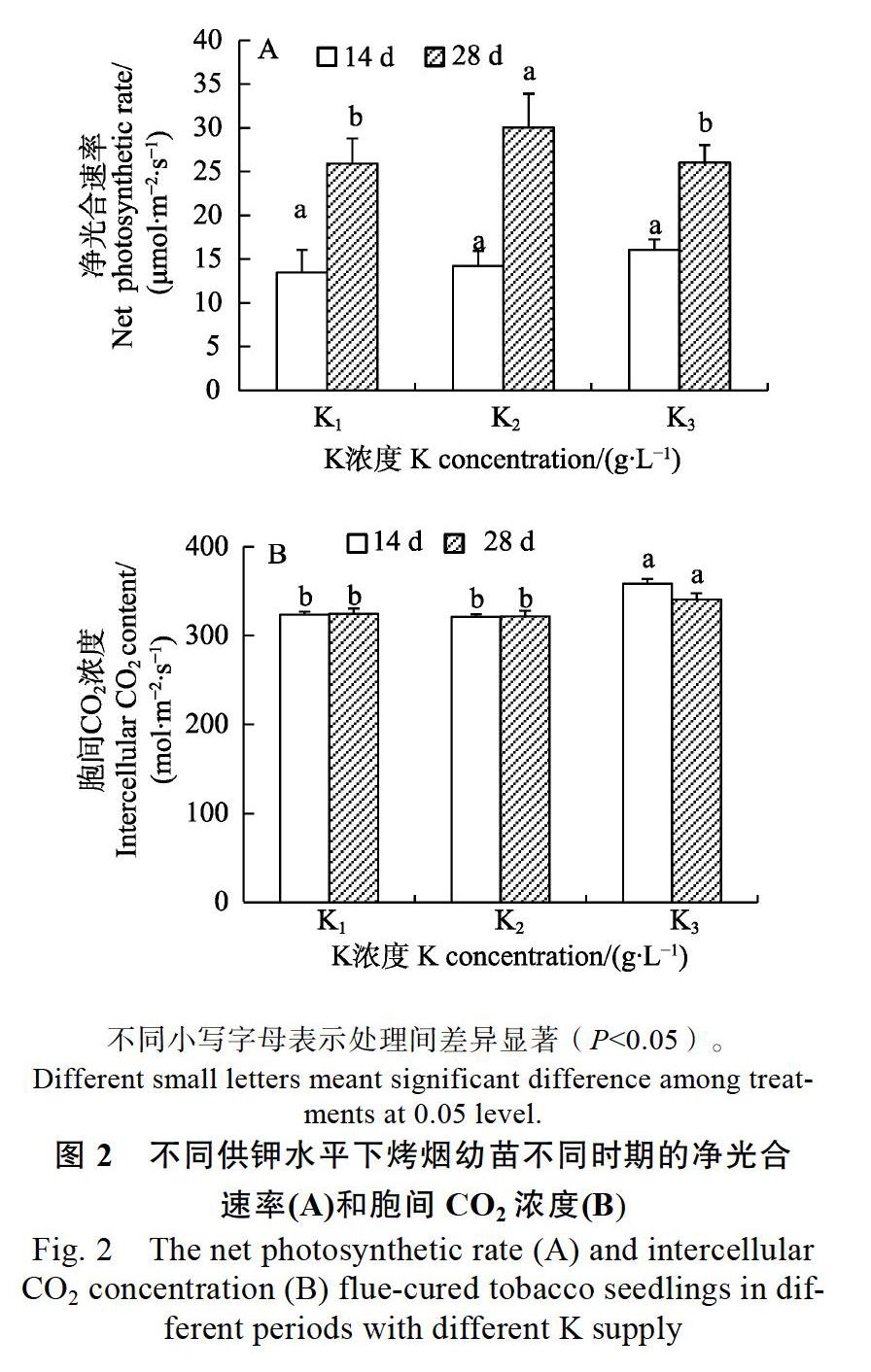
2.3不同施钾量对烤烟净光合速率和胞间CO2浓度的影响
如图2A所示,烤烟幼苗培养至14 d时,不同钾处理间凈光合速率相比差异不显著;培养至28 d时,与低钾(K1)处理相比,中钾(K2)、高钾(K3)处理的烤烟幼苗净光合速率显著增加,而中钾(K2)处理和高钾(K3)处理下的净光合速率差异不显著;与烤烟幼苗培养至14 d相比,培养至28 d的烤烟幼苗净光合速率约提升2倍。结果表明,随着施钾量的提升,对烤烟幼苗前期的净光合速率影响并不显著,但对烤烟生长后期的净光合速率有显著的提升。
如图2B所示,在烤烟幼苗培养至14 d时,与低钾(K1)和中钾(K2)处理相比,高钾(K3)处理的烤烟幼苗的胞间CO2浓度差异极显著,而低钾(K1)处理和中钾(K2)处理下的烤烟幼苗的胞间CO2浓度差异不显著;烤烟幼苗培养至28 d时,与低钾(K1)和中钾(K2)处理相比,高钾(K3)处理的烤烟幼苗的胞间CO2浓度差异显著,而低钾(K1)处理和中钾(K2)处理下的烤烟幼苗的胞间CO2浓度差异不显著。结果表明,随着施钾量的增加,烤烟幼苗的净光合速率及胞间CO2浓度均有提升。但在高钾水平下, 烤烟的各项光合指标增长趋势不显著。
2.4不同钾浓度对烤烟根系生长发育的影响
由图3可知,与低钾(K1)水平和高钾(K3)水平相比较,烤烟幼苗在中钾(K2)水平处理下的根系总根长、总表面积、平均直径和总体积显著增加;低钾(K1)处理与高钾(K3)处理下的烤烟幼苗的根系总根长、总表面积、平均直径和总体积差异不显著。结果表明,适当增加施钾量对根系生长有促进作用,但高钾水平反而抑制根系正常生长。
2.5不同浓度KCl处理对烤烟幼苗硝态氮吸收动力学曲线
由图4可知,烤烟幼苗对NO3--N的吸收速率均随溶液中NO3--N浓度的增加而增大。对高亲和力系统(图4A)来说,所有钾处理下烤烟幼苗的NO3--N吸收曲线均符合Michaelis-Menten方程的描述;中钾(K2)处理烤烟幼苗NO3--N的吸收速率与低钾(K1)处理相比有明显提高;高钾(K3)处理烤烟幼苗NO3--N的吸收速率与低钾(K1)处理相比有明显的降低。可以看出高钾(K3)处理降低了烤烟幼苗NO3--N的吸收速率,而中钾(K2)处理提高了NO3--N的吸收速率,表明烤烟幼苗高亲和力系统受钾浓度影响较大,适宜的钾浓度下烤烟幼苗根系高亲和力系统能够提高对NO3--N的吸收。
对低亲和力系统(图4B)来说,烤烟幼苗对NO3--N的吸收速率均随溶液中NO3--N浓度的增加而增加。中钾(K2)和高钾(K3)处理烤烟幼苗NO3--N的吸收速率与低钾(K1)处理相比有明显提高,但是高钾(K3)处理烤烟幼苗NO3--N的吸收速率比中钾(K2)处理烤烟幼苗NO3--N的吸收速率提高更加迅速。可以看出提高钾水平供应可提高烤烟幼苗NO3--N的吸收速率,表明中钾(K2)和高钾(K3)处理均能提高烤烟幼苗根系低亲和力系统对NO3--N的吸收。
2.6不同浓度KCl处理对烤烟幼苗吸收NO3--N动力学特征的影响
按照Michaelis-Menten方程的Hofstee转换式处理数据,得到不同预处理烤烟幼苗对NO3--N吸收的最大速率(Vmax)和米氏常数(Km)。这2
个参数可用来表征养分离子吸收的动力学过程特点。Vmax表示离子吸收所能达到的最大速率,Vmax越大,离子吸收的内在潜力越大。Km表示根系吸收位点对离子的亲和力大小,Km愈小,亲和力愈大[10]。
由表2可知,无论中钾处理还是高钾处理,与低钾处理相比,烤烟幼苗NO3--N吸收的Vmax均增大,但Km值变化不同。与低钾处理相比,中钾處理烤烟幼苗NO3--N吸收的Vmax增加29.0%,亲和力增加30.0%;K3处理下的烤烟幼苗NO3--N吸收的Vmax增加3.0%,亲和力增加12.0%。与K2处理相比,K3处理下的烤烟幼苗NO3--N吸收的Vmax降低20.0%,亲和力降低21.0%。可以看出施用钾增加了烤烟幼苗吸收NO3--N的最大速率Vmax幅度,同时增加了NO3--N的亲和力。K2处理的Vmax增幅高于K3处理,K2处理的亲和力也明显高于K3处理。
3 讨论
钾是烟草吸收量最大的营养元素,他以离子态被烟株吸收储存于细胞内,具有高速透过生物膜和与酶促反应关系密切的特点[13-15]。钾不仅在生物物理和生化方面具有重要作用,而且对体内同化产物的运输、能量转变也有促进作用,还能促进光合作用,提高CO2的同化率,促进蛋白质的合成[16]。钾也是许多酶(合成酶、氧化还原酶和转移酶)的活化剂,对植物体内C、N代谢作用、细胞代谢、气孔运动具有一定的调节作用[15-16]。钾还能促进光合作用产物向储存器官的运输,增加“库”的贮存,对调节“源”和“库”的相互作用具有良好作用[17]。本研究表明,中钾(K2)水平处理提高烤烟地上部、根系鲜干重,即适当增加钾的施用量有利于增加烤烟地上部和根系的生物量,提高烤烟产量(表1)。施钾能够增加烤烟地上部及根系钾含量;同时在一定钾浓度范围内,烟叶全钾含量与施钾用量存在显著正相关;随钾浓度增加,钾由根系向地上部运输比例增加(图1),而烤烟生育前期净光合速率无显著差异,但后期净光合速率显著增加(图2);中钾(K2)水平增加根系总根长、总表面积、平均直径和总体积,而高钾(K3)水平反而有抑制作用(图3)。
氮与钾是烤烟生长所需的两种主要元素,钾可影响到植物NO3--N的吸收和转运[6-7],缺钾后根系形态发生明显变化,而根系吸收NO3--N的能力与根系形态存在关联[11, 18]。通过利用Epstein建立的植物对养分吸收动力学的研究方法,Vmax表示底物饱和时的反应速度,Km表示当反应速度为最大速度一半时的底物浓度,Km值越小表示不需要很高的底物浓度便可达到最大反应速度,对底物亲和力越大[19]。由于Vmax主要体现离子载体的运转速度,而Km主要体现离子与载体之间的亲和性[20]。本研究表明,经过钾胁迫后,烤烟根系对NO3--N吸收符合离子吸收动力学模型,其吸收动力学参数表现为中、高钾预处理浓度增加后,Vmax增大,但Km值变化不同。施钾水平增加烤烟幼苗吸收NO3--N的最大速率Vmax幅度,同时增加了NO3--N的亲和力。中钾(K2)处理的Vmax增幅高于高钾(K3)处理,中钾(K2)处理的亲和力显著高于高钾(K3)处理。烤烟吸收NO3--N的Vmax与Km值受钾水平的影响很大,供钾水平影响根细胞膜上载体与NO3-之间的亲和性,同时影响载体对NO3-的运转速度。本研究结果显示,经过氮饥饿后,硝态氮的吸收速率增加,这可能与植物体的自身代谢水平及对外界钾环境变化的适应性调节能力密切相关。
NO3-吸收受次生跨膜转运(如载体和离子通道)系统所控制,吸收NO3--N时需要能量,且有ATP酶参加[21-22]。烤烟经过不同浓度的钾预处理后,测定硝态氮的吸收速率,发现与低钾(K1)预处理相比,高钾(K3)预处理降低烤烟根系高亲和力系统对NO3--N的吸收速率,而中钾(K2)处理提高烤烟根系高亲和力系统的NO3--N的吸收速率,表明烤烟根系吸收NO3--N的高亲和力系统受钾水平调控,适宜钾浓度预处理提高烤烟根系对低浓度NO3--N的吸收。对低亲和系统来说,与低钾预处理相比,中钾(K2)和高钾(K3)预处理均提高了烤烟根系低亲和力系统对NO3--N的吸收,表明长期高钾环境生长下的烤烟,根系对硝态氮吸收的低亲和力系统影响较小,而长期低钾环境生长下的烤烟,根系对硝态氮吸收低亲和力系统有抑制作用。
钾是影响烤烟品质的主要元素之一,而且钾与氮的吸收密切相关。目前在我国烟草生产实践中,氮、钾素管理主要存在以下两个问题:一是氮、钾肥投入占肥料总投入的比例逐渐提高,施氮量过大而加重烤烟氮、钾供应的不平衡,容易导致烟叶品质下降,而长期钾肥投入不足及氮肥过高投入,抑制烤烟根系对氮钾吸收,同时也造成氮肥资源的巨大浪费,增加烤烟种植成本;二是改善烤烟产质量的关键取决于土壤供钾能力及氮、钾平衡配施问题,包括基肥氮、钾配方的因土调整和搭配,适当基追氮、钾比例的协调配合和氮肥减量优化施用,采用高效基肥氮、钾配方配合追肥氮、钾平衡供应的技术,有效促进烤烟氮、钾养分供应的协调和平衡,从而协调烤烟碳氮代谢,提高烤烟致香物质的积累水平。
4 结论
不同供钾水平预处理后,烤烟对NO3--N吸收符合离子吸收动力学模型,其吸收动力学参数表现为钾水平预处理浓度增加后,Vmax增大,Km值增加,但增加的幅度不一致。对高亲和系统来说,所有钾预处理烤烟的NO3--N吸收曲线符合Michaelis-Menten方程的描述;与低钾(K1)预处理相比,高钾(K3)预处理降低烤烟根系高亲和力系统对NO3--N的吸收速率,而中钾(K2)处理提高烤烟根系高亲和力系统的NO3--N的吸收速率,表明烤烟根系吸收NO3--N的高亲和力系统受钾水平调控,适宜钾浓度预处理提高烤烟根系对低浓度NO3--N的吸收。对低亲和系统来说,与低钾预处理相比,中钾(K2)和高钾(K3)预处理均提高了烤烟根系低亲和力系统对NO3--N吸收,表明长期高钾环境生长下的烤烟,根系对硝态氮吸收的低亲和力系统影响较小,而长期低钾环境生长下的烤烟,根系对硝态氮吸收低亲和力系统有抑制作用。本研究结果表明,K作为伴随阳离子时会有促进烤烟对NO3--N的吸收和运输;K+是NO3-的主要反離子,当NO3--N吸收和运输到地上部分时,包括K+吸收和运输。此时,高浓度的K更有利于高浓度NO3--N从根部向地上部运输,但抑制了低浓度NO3--N吸收,合适的钾素供应对低浓度NO3--N吸收。但是增加N素供应也可提高地上部K+的含量,从而促进K+的吸收和选择性向上运输,缺K+抑制NO3--N从根系转移到地上部,导致大量NO3--N积累在根部。
参考文献
- 宋国菡, 杨献营, 潘吉焕. 我国烤烟施肥现状、存在问题及对策[J]. 中国烟草科学, 1998(4): 32-34.
- 窦丰科, 张景略. 烟草品质与土壤肥料[M]. 郑州: 河南科学技术出版社, 1992.
- 李絮花, 杨守祥. 施用钾肥对烤烟叶片中钾素和氮素含量的影响[J]. 中国烟草学报, 2002, 8(3): 17-21, 25.
- 高迟銮, 周冀衡, 邱 尧, 等. 氮素和钾素胁迫对去顶烤烟生理指标及烟碱的影响[J]. 作物研究, 2013(5): 449-452.
- 张一扬, 肖汉乾, 李明德, 等. 钾素营养对烤烟生长及养分吸收的影响[J]. 土壤通报, 2004, 35(4): 466-469.
[6] 童依平, 蔡 超, 刘全友, 等. 植物吸收硝态氮的分子生物学进展[J]. 植物营养与肥料学报, 2004, 10(4): 433-440.
[7] 胡青荻. 水稻钾离子转运蛋白基因影响生长发育和硝酸根吸收转运的作用机制[D]. 南京: 南京农业大学, 2016.
[8] 田 歌, 王 芬, 彭 玲, 等. 不同供钾水平对平邑甜茶幼苗生长及NO3-吸收利用特性的影响[J]. 应用生态学报, 2017, 28(7): 2254-2260.
[9] 刘 学, 欧阳由男, 王会民, 等. 水稻根系生长发育特性及其与产量形成的关系[J]. 中国稻米, 2009(4): 8-12.
[10] 丁效东, 张士荣. NaCl对大麦硝态氮吸收动力学特征的影响[J]. 中国生态农业学报, 2015(11): 1423-1428.
[11] Larsson C M, Ingemarsson B. Molecular aspects of nitrate uptake in higher plants[M]//Wary J L, Kinghorn J R. Molecular and Genetic Aspects of Nitrate Assimilation. New York: Oxford University Press, 1989.
猜你喜欢
天津农业科学(2022年6期)2022-07-19
健康体检与管理(2022年4期)2022-05-13
中学化学(2016年10期)2017-01-07
中国高新技术企业(2016年32期)2016-12-27
艺术科技(2016年9期)2016-11-18
中学化学(2015年8期)2015-12-29
第二课堂(课外活动版)(2015年3期)2015-10-21
优雅(2015年9期)2015-09-07
祝您健康(1985年1期)1985-12-29
