
广西斑茅表型性状遗传多样性分析
2019-10-22 01:18黄玉新张保青高轶静段维兴周珊杨翠芳张革民王泽平
热带作物学报 2019年9期
黄玉新 张保青 高轶静 段维兴 周珊 杨翠芳 张革民 王泽平

摘 要 為有效评价和利用广西斑茅种质资源,扩增甘蔗遗传基础。对183份广西斑茅种质资源主要表型性状及遗传多样性进行分析。结果表明:广西斑茅种质资源表型遗传多样性比较低,13个描述型性状的遗传多样性指数在0.0000~1.2349之间,平均为0.3070,以毛群较高,生长带形状较低,空心、气根、根点排列和脱叶性4个性状无多态性表现;不同地区的斑茅资源遗传多样性指数在0.2851~0.5072之间,且以钦州的多样性最大,其次是桂林和崇左,以来宾的多样性最小。5个数值型性状的变异系数在13.54%~29.11%之间,平均为19.59%,以叶宽比较大,叶长较小;10个地区的斑茅资源变异系数在16.48%~21.92%之间,以桂林最大,百色最小。通过聚类分析,183份资源可以分为10个类群,各类群遗传分化不明显,与地理来源无密切联系。本研究揭示了广西不同地区斑茅的表型特异性和遗传多样性,为斑茅资源的采集、保育和杂交利用提供理论参考。
关键词 斑茅;种质资源;表型性状;遗传多样性中图分类号 S566.1 文献标识码 A
Phenotypic Traits and Genetic Diversity ofErianthus arundinaceumGermplasm from Guangxi
HUANG Yuxin, ZHANG Baoqing, GAO Yijing, DUAN Weixing*, ZHOU Shan, YANG Cuifang, ZHANG Gemin, WANG Zeping
Sugarcane Research Institute, Guangxi Academy of Agricultural Sciences / Sugarcane Research Center, Chinese Academy of Agricultural Sciences / Guangxi Key Laboratory of Sugarcane Genetic Improvement / Key Laboratory of Sugarcane Biotechnology and Genetic Improvement (Guangxi), Ministry of Agriculture and Rural Affairs, Nanning, Guangxi 530007, China
Abstract This study was conducted to effectively evaluate and utilize the germplasm resources ofErianthusarundinaceumin Guangxi, to expand the genetic basis of sugarcane. The phenotypic traits and genetic diversity of 183 accessions ofE. arundinaceumcollected from Guangxi were analyzed. Results showed that the genetic diversity of phenotypic traits was low. The genetic diversity index (DI) ranged of 13 descriptive traits from 0.0000 to 1.2349 with an average of 0.3070, with highest for hair groups and lowest for growth ring shape. There was no polymorphicin 4 traits including pipe, aerial root, root primordial and sheath detached. The DI among different regions ranged from 0.2851 to 0.5072, with highest for Qinzhou, followed by Guilin and Chongzuo, and the lowest for Laibin. The coefficient of variation (CV) of 5 numerical traits ranged from 13.54% to 29.11% with an average of 19.59%, with highest for leaf width and lowest for leaf length. The regional CV of 10 regions ranged from 16.48% to 21.92%, with highest for Guilin and the lowest for Baise. Cluster analysis showed that 183 germplasm resources could be clustered into 10 groups. The genetic differentiation among groups was not obvious and had no close relationship with geographical origin and the genetic distance was not closely related to geographical origin. This study revealed the phenotypic specificity and genetic diversity ofE. arundinaceain different regions of Guangxi, and provided theoretical reference for collection, conservation and hybridization ofE. arundinacea resources.
Keywords Erianthus arundinacea; germplasm resources; phenotypic traits; genetic diversity
DOI10.3969/j.issn.1000-2561.2019.09.006
遗传多样性是指种内不同种群之间或一个种群内不同个体之间的遗传差异,多样化的遗传资源是品种改良的基础。由于长期的近亲杂交和高产、高糖等有益性状的定向选择,现代甘蔗品种遗传力逐渐趋于一致,因此,甘蔗野生种属血缘的渗入,是提高甘蔗异质性,扩大遗传变异的重要途径。斑茅(Erianthus arundinaceum)是甘蔗近缘属植物之一,其抗逆和抗病虫性强、适应性广、丛生性好、分蘖多、生势强、宿根性好、生态类型丰富,对甘蔗品种改良具有重要的应用潜力。前人对斑茅在抗寒[1]、抗旱[2-4]、抗病虫害[5-6]、杂交利用等[7-9]方面进行了较多研究;在遗传多样性方面,蔡青[10]通过花粉母细胞观察,斑茅染色体有2n=30、40、60等3种类型;张木清等[11]、徐荣等[12]、张建波等[13]和宋焕忠等[14]分别利用ISSR、SARP和SCoT分子标记对斑茅种质遗传多样性进行研究,研究结果均显示,供试材料呈现出较强的地域分布,材料间的遗传距离与地理距离有一定的相关性;梁緒振等[15]研究结果揭示了四川地区斑茅具有较为丰富的遗传多样性;徐超华等[16]研究结果表明,全国9个省162份斑茅种质质量性状Shannon-Wiener 多样性指数整体偏低,数量性状的遗传变异较质量性状丰富。种质资源是育种的基础,国内外育种单位都非常重视甘蔗种质资源的收集和保存工作。目前,广西甘蔗种质资源圃保育有一批采自广西不同地区(市、县)的斑茅种质,但对于这些种质材料的遗传多样性水平和不同地区群体间的异同点尚缺乏足够的认识。本研究通过调查5个数量性状和13个质量性状对183份广西斑茅进行表型多样性评价分析,旨在了解广西斑茅种质的遗传多样性和变异情况,为斑茅资源的采集、保育和杂交利用提供理论参考。
1 材料与方法
1.1材料
以广西甘蔗种质资源圃内保育的183份广西斑茅种质资源为研究材料。按采集地点统计,桂林市34份(编号GXE1~GXE34),柳州市25份(编号GXE35~GXE59),来宾市14份(编号GXE60~GXE73),贵港市13份(编号GXE74~ GXE86),玉林市9份(编号GXE87~GXE95),崇左市14份(编号GXE96~GXE109),钦州市8份(编号GXE110~GXE117),南宁市18份(编号GXE118~GXE135),河池市28份(编号GXE136~GXE163),百色市20份(编号GXE164~GXE183)。
1.2 方法
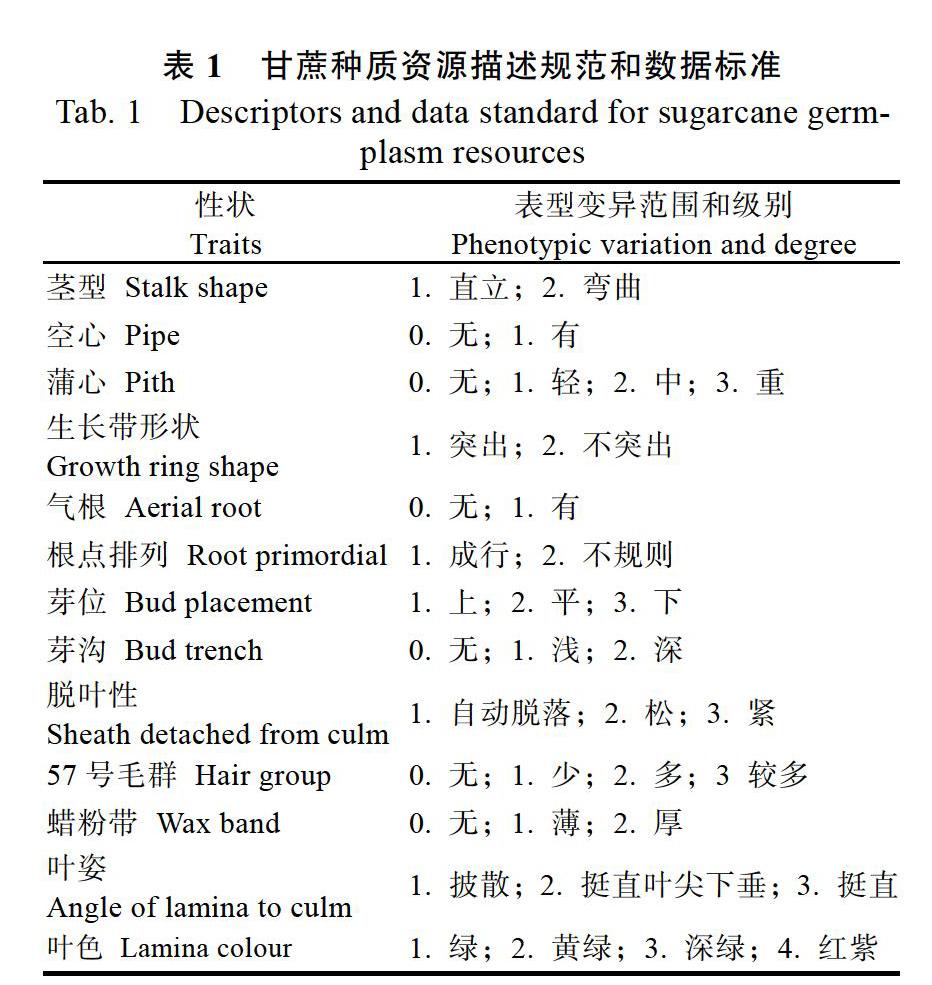
试验于2016年11月在广西甘蔗种质资源圃进行。所有材料均保育在直径0.7 m、框距1.2 m的水泥框里。选择5个数量性状(株高、茎径、叶长、叶宽、锤度)和13个质量性状(茎形、空心、蒲心、生长带形状、气根、根点排列、芽位、芽沟、脱叶性、57号毛群、蜡粉带、叶姿、叶色)进行调查,并从中选取9个有多样性表现的质量性状(茎形、蒲心、生长带形状、芽位、芽沟、57号毛群、蜡粉带、叶姿、叶色)进行广西斑茅种质表型遗传多样性分析。所有描述型和数值型性状均参照《甘蔗种质资源描述规范和数据标准》[17](表1)进行观测和测定。
1.3数据处理
利用SPSS 17.0软件对数量性状的变异情况进行分析,采用Shannon-Wiener多样性指数(Shannon-Wiener diversity index,H′)进行遗传多样性评价,其计算公式H′=-ΣPiLnPi,式中Pi为某性状第i个代码值出现的频率,Ln表示自然对数[18]。参考吴建涛等[19]方法利用模糊隶属函数计算出数量性状的隶属函数值使用DPS18.10进行聚类分析,其中聚类方法采用类平均法(UPGMA),种质间遗传距离为卡方距离。
2结果与分析
2.1质量性状的遗传多样性
由表2可知,183份广西斑茅质量性状的Shannon-Wiener多样性指数在0.0000~1.2349之间,平均值为0.3070,总体偏低。其中以毛群的多样性最高(1.2349),多样性较丰富;其次是叶色(0.8550)、叶姿(0.6913)和蜡粉带(0.5645),多样性中等;茎型、蒲心、生长带形状、芽位和芽沟多样性水平较低;空心、气根、根点排列和脱叶性4个性状无多态性表现。总体而言,广西斑茅的类型主要为无气根,根点成型排列,难脱叶,大部分材料表现为直茎(94.5%),重度蒲心(94.0%)、生长带形状不突出(99.5%),平位芽(98.4%)、无芽沟(98.9%),蜡粉带较薄(77.0%),叶片以黄绿色(71.0%)为主,叶姿挺直(53.0%)或挺直叶尖下垂(47.0%),除少数材料无57号毛群外,其余大部分材料都有,其中38.8%毛群少,21.9%毛群多,33.3%毛群较多。
2.2数量性状的变异情况
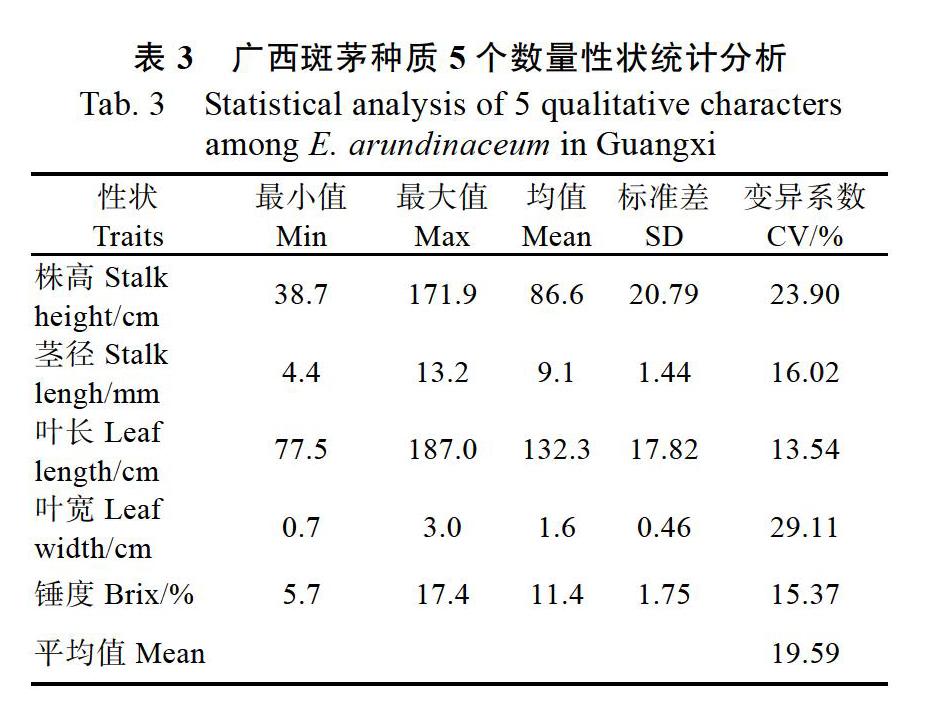
对5个数量性状变异情况进行统计分析,由表3可知,5个数量性状的变异系数在13.5%~29.1%之间,平均为19.6%。各个性状的变异程度依次为叶宽(29.1%)>株高(23.9%)>茎径(16%)>锤度(15.4%)>叶长(13.5%)。因此,广西斑茅数量性状的主要变异来源于叶宽,其次是株高,茎径、叶长和锤度的变异差异不大。
2.3 不同地区斑茅种质资源的表型遗传差异
2.3.1 质量性状的遗传差异 对广西不同地区斑茅种质资源的遗传多样性进行比较(表4),其中除贵港斑茅以叶色多样性(0.9372)最高外,其他9个地区斑茅均以57号毛群的多样性最高。而生长带形状在桂林斑茅存在多样性外,其他9个地区斑茅的生长带形状均无多态性;芽位和芽沟两个性状在70%的地区也无多态性。10个地区斑茅平均多样性指数在0.2851~0.5072之间,且以钦州的多样性最大,来宾的多样性最小,其次依次为桂林(0.4523)>崇左(0.4228)>百色(0.4126)>河池(0.4066)>柳州(0.3635)>南宁(0.3534)>玉林(0.3238)>贵港(0.2968)。
2.3.2 数量性状的遗传差异 对不同采集地区斑茅种质资源的5个数量性状比较可知(表5),崇左斑茅平均株高(104.2 cm)、平均茎径(10.0 mm)和平均叶长(142.1 cm)都具有最大值;玉林斑茅平均叶宽(2.0)最大,钦州斑茅平均锤度(12.0%)最高;而南宁斑茅的株高(71.3 cm)和茎径(8.1 mm)最小,玉林的叶长(126.6 cm)最短,来宾的叶宽(1.3 cm)最窄,玉林的锤度(10.2%)最低。从不同地区5个数量性状的变异系数来看,株高的变异系数以钦州(35.46%)的最大,百色的(14.95%)最小;茎径的变异系数也以钦州(22.83%)的最大,河池(11.01%)的最小;叶长的变异系数以桂林(19.29%)的最大,钦州(8.92%)的最小;叶宽的变异系数以桂林(34.03%)的最大,玉林(16.53%)的最小;锤度的变异系数以玉林(22.15%)的最大,钦州的最小(6.27%)。
整体而言,桂林斑茅的叶长和叶宽具有较大的遗传差异;柳州斑茅5个性状平均值与桂林斑茅差异不大,除锤度外,其他4个性状的变异程度都比桂林斑茅低;来宾斑茅叶宽最小,锤度也较低;贵港斑茅各性状处于中等水平;玉林斑茅叶片短、宽,锤度虽变异系数大,但总体偏低;崇左斑茅植株高大,叶子即长又宽;钦州斑茅锤度高,株高和茎径具有较大的遗传差异;南宁斑茅较矮小,茎径较细;河池斑茅茎径较粗;百色斑茅平均茎径和锤度均排行第二。
2.4聚类分析
根据广西斑茅资源的表型性状(5个数量性状和9个质量性状)进行聚类分析(图1),所有参试材料在遗传距离1.35处可分为10类。第Ⅰ类和第Ⅳ类分别有109和40个材料,均来自10个不同地区;第Ⅱ类有19个材料,来自除玉林和崇左外的8个地区;第Ⅲ类有2个材料,GXE4和GXE6,均来自桂林;第Ⅴ类有2个材料,GXE12和GXE60,分别来自桂林和来宾;第Ⅵ类有5个材料,来自4个不同地区;第Ⅶ类有2个材料,GXE104和GXE106,均来自崇左;第Ⅷ类有2个材料,分别来自钦州(GXE113)和南宁(GXE125);第Ⅸ类和第Ⅹ类都只有1个材料,分别是GXE121和GXE182。从聚类结果来看,材料的划分与地理来源无明显的联系,但也有少部分来源相同的材料优先聚在一起,如第Ⅲ类和第Ⅶ类,来自相同地区的2个材料聚为一类。同时,值得注意的是第Ⅸ类和第Ⅹ类均只有一个材料,与其他类群分開。
3 讨论
对作物种质资源的遗传多样性进行调查和分析,是种质资源研究的首要工作,也是作物育种的基础性工作。近年来,国内外学者对甘蔗品种或甘蔗近缘种属表型性状或分子水平的遗传多样性进行了一些研究,为甘蔗遗传育种亲本选择、杂交组合配制以及甘蔗野生资源的采集、保育和创新利用提供了有效的参考[19-26]。本研究对广西省内不同地区(市)斑茅表型差异比较发现,在崇左市容易采集到高大型斑茅,在南宁市容易采集到矮小型斑茅;在桂林市容易采集到数量性状变异大的斑茅,在钦州市容易采集到质量性状多样化的斑茅。
本研究中广西斑茅种质资源表型性状整体偏低,与徐超华等[16]的研究结果基本相同;而宋焕忠等[14]研究发现广西斑茅具有较丰富的遗传多样性,本研究结果与其研究结果不同,原因可能是(1)本研究调查的性状只是表型性状的一部分,不能全面代表广西斑茅的表型变异;(2)本研究是基于表型性状数据的分析,而宋焕忠等[14]研究是从分子水平进行的多样性评价,表型性状是基因和环境共同作用的结果,可能有些发生在DNA水平的变异不一定造成表型性状的改变。薛德等[27]对五节芒表型性状和SSR标记遗传多样性分析,也发现表型性状数据的聚类结果与SSR标记聚类结果并不完全一致。聚类分析结果表明,183份广西斑茅遗传分化不明显,供试材料的聚类结果与地理来源没有严格的一致性,与刘新龙等[25]和刘建乐等[26]的研究结果基本相同。笔者认为广西各地区斑茅遗传分化不明显的原因可能是斑茅主要以风为媒介,依靠种子进行传播,而广西省内河流较多(如桂林漓江,柳州柳江,百色右江,河池龙江,崇左左江,贵港郁江,南宁邕江等),各河流纵横交错,促进了不同流域的基因交流,使得各地区的材料遗传差异不明显。值得注意的是,第Ⅸ类群采自南宁的斑茅GXE121以及第Ⅹ类群百色斑茅GXE182均单独聚为一类,与其他类群分开,值得重点关注。
前人研究发现作物的表型变异与其生境有一定的相關性[28-29]。徐超华等[16]研究结果表明,斑茅的锤度与海拔高度呈极显著负相关,株高与纬度呈极显著负相关。刘新龙等[25]研究发现,国内十倍体割手密资源的数量性状与采集地的海拔、纬度也具有相关性,其中产量性状与纬度和海拔呈负相关;糖分性状与海拔呈负相关,与纬度呈正相关;纤维分与海拔呈正相关。本研究的试验材料大部分都是早期采集,可能由于当时没有系统的采集计划,同时缺乏相应的采集设备,对海拔、经纬度等地理信息没有进行全面地收集,因此,无法对本研究的材料与地理环境的关系进行分析。目前,广西种质资源圃内大约保育有300多份广西斑茅资源,大约1/2的资源具有较详细的地理信息数据。这些采集信息比较全面的资源可反映广西斑茅资源分布情况,也可用于数量性状与地理环境的关系研究。
甘蔗野生种质资源是未来优质甘蔗品种选育的基础,如何良好地利用这些种质资源为甘蔗新品种的有效选育提供条件,还需要找出一个更合理有效的方法。本研究虽然包含了广西10个地区(市)的材料,但各地区的材料主要集中在该地区的少数几个县,大部分县的材料都没有;同时还缺乏广西另外4个地区(贺州市、防城港市、北海市和梧州市)的试验材料。因此,要对广西斑茅类型和遗传多样性情况进行更深入的了解,获得更为全面的试验结果,需扩大试验群体,增加更多不同地区(市、县)的斑茅样本量,同时结合形态标记和分子标记等方法进行研究。
参考文献
- Ram B, Sreenivasan T V, Sahi B K, et al. Introgression of low temperature tolerance and red rot resistance from Erianthus in sugarcane[J]. Euphytica, 2001, 122(1): 145-153.
- 陈义强. 甘蔗抗旱种质资源的筛选及斑茅杂种后代抗旱性分析[D]. 福州: 福建农林大学, 2005.
- 吴水金, 潘世明, 陈义强, 等. 甘蔗斑茅属间远缘杂种真实性的ITS鉴定及其抗旱性分析[J]. 江西农业大学学报, 2008, 30(4): 628-632.
- 黄忠兴, 符 成, 黄锦福, 等. 含斑茅血缘甘蔗亲本的抗旱性研究[J]. 广东农业科学, 2016, 43(6): 49-57.
- Piperidis G, Christopher M J, Carroll B J, et al. Molecular contribution to selection of intergeneric hybrids between sugarcane and the wild species Erianthus arundinaceus[J]. Genome, 2000, 43(6): 1033-1037.
- 李文凤, 蔡 青, 黄应昆, 等. 甘蔗野生资源对蔗茅柄锈菌的抗性鉴定[J]. 植物保护, 2005, 31(2): 51-53.
- 王丽萍, 蔡 青, 范源洪, 等. 甘蔗(Saccharum)与斑茅(Erianthus arundinaceus)远缘杂交利用研究[J]. 西南农业学报, 2007, 20(4): 721-726.
- Jackson P, Henry R J. Wild crop relatives genomic & breeding resources[M]. Berlin: Springer, 2011: 97-107.
- Piperidis N, Chen J W, Deng H H, et al. GISH characterization of Erianthus arundinaceus chromosomes in three generations of sugarcane intergeneric hybrids[J]. Genome, 2010, 53(5): 331-336.
- 蔡 青. 甘蔗近缘属斑茅、滇蔗茅遗传多样性与种质创新研究[D]. 昆明: 云南大学, 2013.
- 张木清, 洪艺珣, 李奇伟, 等. 中国斑茅种质资源分子多态性分析[J]. 植物资源与环境学报, 2004, 13(1): 1-6.
- 徐 荣, 何丽莲, 王先宏, 等. 斑茅野生种质资源遗传多样性的ISSR分析[J]. 分子植物育种, 2017, 15(7): 2874-2880.
- 张建波, 鄢家俊, 张蕴薇, 等. 斑茅野生种质资源SRAP遗传多样性研究[J]. 生物技术学报, 2013, 21(10): 1193-1202.
- 宋焕忠, 张荣华, 杨海霞, 等. 广西斑茅遗传多样性的SCoT标记分析[J]. 西南农业学报, 2014, 27(1): 59-64.
[15] 梁绪振, 鄢家俊, 白史且, 等. 斑茅种质资源形态性状的变异研究[J]. 草业科学, 2011, 28(7): 1307-1314.
[16] 徐超华, 陆 鑫, 马 丽, 等. 斑茅种质资源的表型性状及遗传多样性[J]. 湖南农业大学学报(自然科学版), 2014, 40(2): 117-121.
[17] 蔡 青, 范源洪. 甘蔗种质资源描述规范和数据标准[M]. 北京: 中国农业出版社, 2006: 7-31.
[18] 孔繁玲. 植物数量遗传学[M]. 北京: 中国农业大学出版社, 2006.
[19] 吴建涛, 许环映, 谢 静, 等. 粤糖系列甘蔗亲本表型性状遗传多样性分析[J]. 植物遗传资源学报, 2018, 19(4): 748-759.
[20] Singh P, Singh S P, Tiwari A K,et al. Genetic diversity of sugarcane hybrid cultivars by RAPD markers[J]. Biotech, 2017, 7(3): 222.
[21] 胡 杨, 李 赟, 黄有总, 等. 利用SSR与RAPD分子标记评估甘蔗品种的遗传多样性[J]. 基因组学与应用生物学, 2016, 35(9): 2494-2503.
[22] Oliveira L A R, Machado C A, Cardoso M N,et al. Genetic diversity ofSaccharum complex using ISSR markers[J]. Genetics and Molecular Research, 2017, 16(3): 1-9.
[23] 刘新龙, 蔡 青, 吴才文, 等. 甘蔗品种资源的表型遗传多样性[J]. 生物多样性, 2010, 18(1): 37-43.
[24] Govindaraj P, Amalraj V A, Mohanraj K,et al. Collection, characterization and phenotypic diversity ofSaccharum spontaneumL. from arid and semi arid zones of northwestern India[J]. Sugar Techenology, 2014, 16(1): 36-43.
[25] 刘新龙, 苏火生, 应雄美, 等. 中国十倍体割手密资源的表型相关性及遗传多样性[J]. 湖南农业大学学报(自然科学版), 2012, 38(6): 574-579.
[26] 刘建乐, 白昌军, 严琳玲, 等. 43份割手密资源农艺性状遗传多样性评价[J]. 热带作物学报, 2015, 36(2): 229-236.
[27] 薛 德, 肖 亮, 艾 辛, 等. 五節芒表型性状和SSR标记遗传多样性分析[J]. 草业学报, 2012, 21(5): 96-106.
[28] 杨晓霞, 冷平生, 郑 健, 等. 暴马丁香不同种源种子和幼苗的表型性状变异及其与地理-气候因子的相关性[J]. 植物资源与环境学报, 2016, 25(3): 80-89.
[29] 肖海峻, 徐 柱, 李临杭, 等. 鹅观草表型性状变异与生境间的相关性[J]. 中国草地学报, 2007, 29(5): 22-30.
猜你喜欢
今日农业(2022年5期)2022-07-02
安徽农业科学(2022年9期)2022-05-17
安徽农学通报(2022年6期)2022-04-07
瞭望东方周刊(2021年6期)2021-03-30
瞭望东方周刊(2021年6期)2021-03-30
种子科技(2020年15期)2020-10-20
三农资讯半月报(2020年2期)2020-03-09
山东农业科学(2019年12期)2019-02-10
农民致富之友(2017年19期)2017-10-21
中国医药导报(2017年13期)2017-07-03
