
施肥对贵南县轻度退化草甸植被特征的影响
2019-09-25 11:03贾志锋雷生春徐成体魏小星刘文辉
草地学报 2019年4期
贾志锋, 马 祥, 雷生春, 徐成体, 魏小星, 刘文辉
(青海大学畜牧兽医科学院/青海省青藏高原优良牧草种质资源利用重点实验室, 青海 西宁810016)
贵南县位于青海省东北部,隶属海南藏族自治州,地处西倾山与黄河之间,属共和盆地的一部分,总面积为6 649.7 km2,境内平均海拔3 100 m,属高原大陆性气候,冬长夏短,有可利用草地面积约40万hm2,耕地面积约2万hm2[1]。畜牧业是贵南县主要经济支柱,境内光、温、水等农牧业气候资源丰富,草质柔软,牧草生长期一般在160~180 d之间,山区牧草生长期少于160 d,是青海省发展牧业生产条件较好的地区之一[2]。
但是,近年来由于自然条件制约、全球变暖和传统观念等的综合作用,该县畜牧业仍以天然草地放牧为主,单纯追求牲畜数量增长,从而导致草地退化、沙化、盐碱化面积不断扩大,草地质量及牧草品质逐年下降,使贵南县草地畜牧业和生态环境等受到很大影响[2]。草地退化表现为植被盖度降低、秃斑面积增大、地上生物量比例降低、可食牧草比例和高度降低、草地生物多样性减少、草地土壤有机质含量降低、土壤质量变劣等多个方面。草地退化程度加剧导致水土流失严重、草地植物群落多样性急剧减少、濒危植物种类不断增加、生态系统稳定性减弱、草地生产力不断降低等严重影响高寒草地植物群落结构及物种多样性的众多问题[3],严重阻碍草地畜牧业的可持续稳定发展和当地牧民生活水平的提高。因此,草地退化问题已引起生态学家、当地牧民及相关政府部门的关注。杨晓玫等[4]研究表明天祝高寒草地轻度退化草地(Light Degradation,LDG)、中度退化草地(Moderate Degradation,MDG)和重度退化草地(Severe Degradation,SDG)植物群落四度一量特征整体呈现逐渐降低趋势且差异显著(P<0.05),表现为LDG> MDG> SDG;随着天祝高寒草地退化程度的增加,其植物多样性逐渐降低,植物群落结构分布单一。草地退化后植被优势种物种数减少,杂类草增加,植被盖度及生物量降低,严重影响了当地畜牧业发展[5]。杨娇等[6]研究表明随着退化程度的增加,紫花针茅(Stipacapillata)种群高度和个体生物量显著降低(P<0.05),分蘖节和茎的直径呈现增加的趋势,而根长、茎长、叶长、花序长呈现减少的趋势。草地退化还可衍生鼠害、虫害及毒杂草泛滥,进一步加剧了草地的退化,加之气候变化(全球变暖、降雨变化等)的影响,使得高寒草地的质量严重下降,草地原生植被减少,其生态价值基本丧失,进一步退化形成黑土滩。从上个世纪70年代到现在,黑土滩成为青海省三江源地区的主要生态问题,严重阻碍了该地区草地畜牧业的可持续发展与利用[7]。贵南县退化草场面积达15.8万hm2,主要分布在牧业三乡的夏秋草场,为毒杂草型、黑土滩型和沙漠化型退化草地[8]。鉴于此,在该县加大退化草地的恢复研究意义重大。根据赵新全[9-10]、董全民[11]、潘多峰[12]对草地退化现状及其退化类型的研究结果,比对观测到的植被盖度(60%~80%)、地上生物量比例(50%~75%)、可食牧草比例(50%~75%)、可食牧草高度(下降 3~5 cm)、秃斑地比例、植被覆盖率[13]和优势植物种类等草地表现,结合GB 19377-2003[14]判断该地区的退化草地类型为轻度退化草甸草地。
施肥是改良退化草地的常用措施,王伟等[15]对高寒草甸进行施氮研究,发现硫酸铵形态的氮肥对地上生物量的增加最为显著,尿素次之,200 kg·hm-2为最佳硫酸铵氮肥施用量。丁洋[16]研究发现施有益微生物菌肥可显著增加草原3号杂花苜蓿(MedicagovariaL. ‘Caoyuan No. 3’)、草原2号杂花苜蓿(MedicagovariaL. ‘Caoyuan No.2’)、蒙农杂种冰草(Agropyroncristatum×A.desertorum‘Mengnong’)、无芒雀麦(Bromusinermis)的干草产量和种子产量,同时可明显改善株丛结构。珊丹等[17-18]研究认为微生物菌肥添加使草原矿区排土场土壤微生物菌落数量增加、土壤酶活性增强,紫花苜蓿(Medicagosativa)和披碱草(Elymusdahuricus)地上生物量显著增加,对增加土壤肥力、加快植被重建有重要作用,沟施法+0.4 mL·m-2菌肥为最佳施肥措施。可见,施肥对草地恢复的影响与草地类型、肥料种类、施肥措施和施肥量密切相关,但贵南县轻度退化草地在施肥改良方面的研究尚未见报道。
因此,本研究在贵南县轻度退化草地进行施肥试验,旨在明晰施肥对退化草地植被群落盖度、高度及生物量的影响,以探讨贵南县轻度退化草地改良的合理施肥方案,为贵南县退化草地生态系统的恢复与重建提供科学依据。
1 材料与方法
1.1 试验地概况
试验地位于青海省贵南县森多乡茫什多滩,海拔3 670 m,东经101°18′,北纬35°46′,年均气温2.3℃,年均降水量400~500 mm,无霜期40 d,≥5℃积温1 500℃~1 800℃。土壤为高山草甸土,土壤pH值为7.0,土壤有机质25.8 g·kg-1。草地类型为高寒草甸,主要优势种有:矮生嵩草(Kobresiahumilis)、垂穗披碱草(Elymusnutans)、溚草(Koeleriacristata)和紫花针茅(Stipacapillata);毒杂草有黄花棘豆(Goxytropisochrocephala)、狼毒(Stellerachamae)和高山唐松草(Thalictrumalpinum)。试验区面积6 666.7 m2,在植被恢复前期进行了地面鼠和地下鼠防治工作。
1.2 试验设计
试验设置7个处理,包括5个施氮处理(60 kg·hm-2,105 kg·hm-2,150 kg·hm-2,195 kg·hm-2,240 kg·hm-2,分别记作N1,N2,N3,N4,N5)和1个菌肥处理(记作J),菌肥为EM菌液(Effective Microorganisus,EM)发酵的牛粪,EM菌液由青海省牧业科技开发公司生产提供,参考雷生春等[19]的研究结果和生产实践中的用量确定施用量为45 m3·hm-2,设置1个对照,氮肥和菌肥均不施,记作N0。于2011年6月10日对轻度退化草地进行施肥处理,肥料选用尿素(46.6% N)和生物有机肥(活菌数≥0.2亿·g-1),均匀撒播,施肥完成同时进行围栏和鹰架(巢)建设。采用随机区组试验设计,区组间距20 m,小区面积225 m2(15 m ×15 m),相邻小区之间设2 m缓冲带,每个处理重复3次。2011年7—10月、2012 年7—10月的每月中旬,对施肥草地和对照进行植被盖度、草群高度、地上生物量的监测。每个样地内按蛇形法随机重复取样6次,样方面积1 m2(1 m × 1 m)。
1.3 指标测定
盖度用针刺法测定,每样方100针;高度用钢卷尺测定,各样方中按经济类群“禾草类”、“莎草类”和“杂草类”每个类群随机选取5株,测其自然高度,群落高度为各类群株高的平均值;每个样方调查完毕后,将样方内各类草齐地剪下后,按经济类群“禾草类”、“莎草类”和“杂草类”进行分离,分别装袋,带回实验室,烘箱105℃杀青,然后烘干至恒重后称重,总生物量为3个经济类群生物量的总和。
1.4 数据处理
采用Excel 2010对数据进行初步整理,采用SPSS 17.0软件进行数据分析。年份与施肥处理间的交互作用对测定指标的影响采用多因子方差分析(Two-way ANOVA),并用Duncan法对施肥处理间数据进行多重比较(P<0.05)。结果用平均数表示,数据误差用均值的标准误(Standard error of mean,SEM)表示。
2 结果与分析
2.1 年份及施肥处理对退化草地恢复过程中主要指标的影响
在施肥处理后不同时间下,年份、施肥处理及两者的互作效应对草地群落盖度、高度及生物量的影响均较大(表1)。2011—2012年,7—10月,除施肥处理对7月群落高度的影响达显著(P<0.05)水平外,其他指标受年份、施肥处理及两者的互作效应的影响均达极显著(P<0.01)水平。
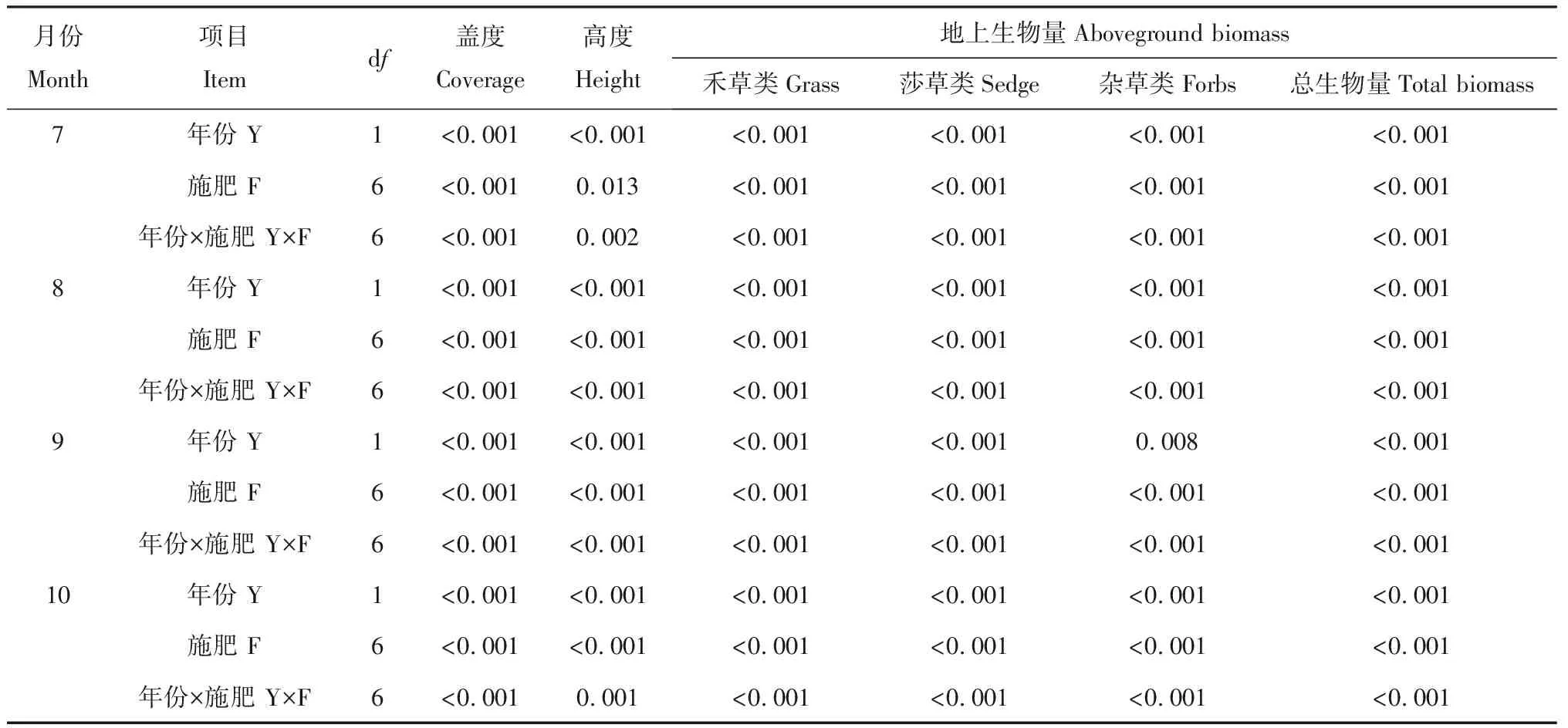
表1 年份及施肥处理交互作用下不同月份各指标的P值Table 1 P value of each factor under different year and fertilization treatment on different month
注:年份:Year,Y;施肥:Fertilization,F
2.2 年份及施肥处理对轻度退化草地盖度的影响
年份及施肥处理均对轻度退化草地盖度有显著影响(表1,图1)。与2011年相比,2012年各施肥处理草地盖度在7—10月均显著(P<0.05)高于2011年;N0组在7—10月也显著(P<0.05)高于2011年,分别高出145.7%,82.8%,27.7%和294.5%。整体而言(表2),2012年各处理的草地盖度均值在7—10月比2011年分别高出79.9%,99.2%,31.2%和271.6%。可见,施肥后第2年退化草地群落盖度在牧草生长期(7—8月)、盛草期(9月)和枯草期(10月)均大幅提高(P<0.05)。
对各施肥处理来说,2011年群落盖度整体有一定差异,但规律性不强。7月时,N4、N5和J施肥处理下,群落盖度显著(P<0.05)高于N0和其它3种低氮处理,即N1,N2和N3。8—10月,各处理间草地群落盖度均有波动,8月、9月和10月群落盖度最高的处理分别为J,N2和N1,较对照分别显著(P<0.05)增加33.3%,21.8%和57.4%。2012年7月时,N4处理群落盖度显著高于对照(P<0.05),较对照高出35.2%,8—10月各施肥处理草地群落盖度均显著高于N0(P<0.05)。N4处理的群落盖度在7—10月均处于较高水平,且随牧草生育期的延长呈上升趋势,10月达到最高(99.3%)。N1,N2,N3,N5和J处理群落盖度在维持较高水平时伴随一定的波动,规律不明显。
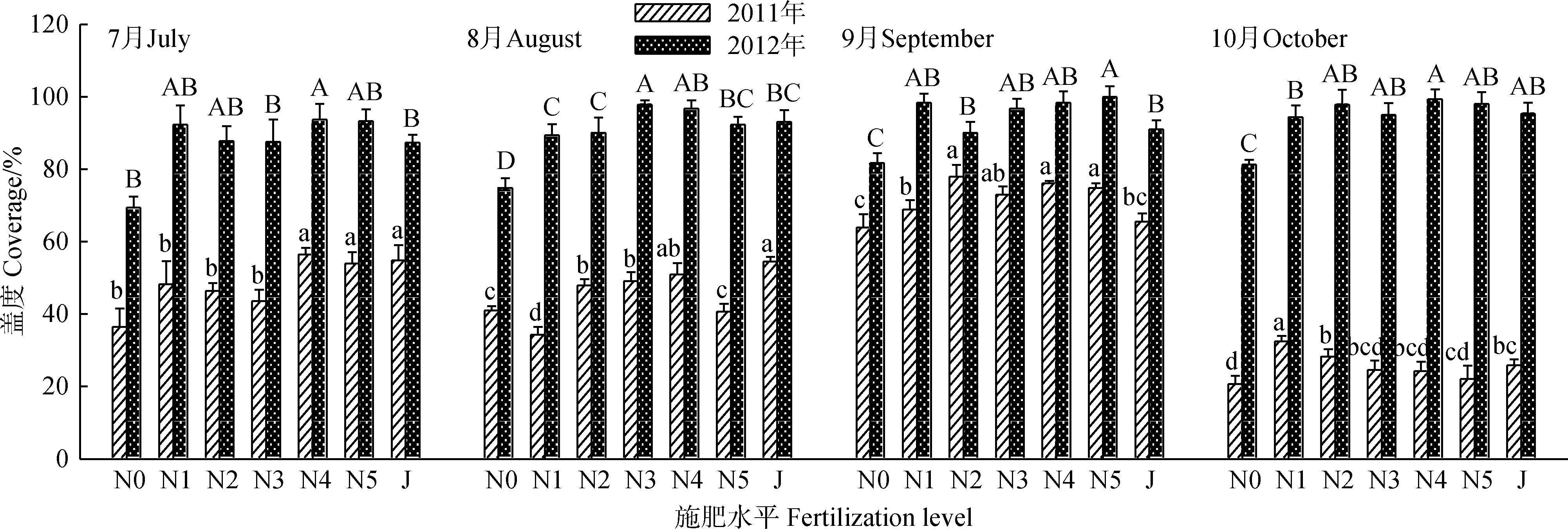
图1 施肥处理对轻度退化草地群落盖度动态的影响Fig.1 Effects of fertilization on vegetation coverage of light degraded meadow注:不同小写字母表示2011年同一月份不同施肥处理间差异显著(P < 0.05),不同大写字母表示2012年同一月份不同施肥处理间差异显著(P < 0.05),下同Note:Different lowercase letters for the same month indicate significant difference among the different years and treatments at 0.05 level in 2011,different uppercase letters for the same month indicate significant difference among the different years and treatments at 0.05 level the in 2012. The same as below
表2 不同处理盖度均值Table 2 Mean coverage of different treatment/%

年份Year月份Month7月July8月August9月September10月October201148.5b45.4b71.4b25.4b201287.3a90.5a93.7a94.4a
注:表中不同小写字母表示差异显著(P< 0.05),下同
Note:Different lowercase letters indicate significant difference at 0.05. The same as below
2.3 年份及施肥处理对轻度退化草地群落高度的影响
年份及施肥处理对轻度退化草地群落高度的影响均较大(表1,图2)。从年份上来看(表3),2012年各施肥处理草地群落高度均显著高于2011年,7—10月平均高度分别比2011年高出142.2%,91.1%,59.7%和204.9%(P<0.05)。2012年N0处理草地群落高度在7月、8月和10月也显著(P<0.05)高于2011年N0处理。
随牧草生育期的延长,2011年各处理的群落高度总体呈现先上升后降低的规律,最大高度集中在8月和9月;2012年则不同,N1、N2和N3处理群落高度总体呈先升后降再升的规律,N4、N5、J和N0处理始终表现为上升,且所有处理的群落高度最大值均在10月。2011年7月和10月,各处理间差异不显著(P>0.05),8月N5和J处理的群落高度较高,显著(P<0.05)高于其它各处理,9月时N0处理的群落高度高于N1,N3和N5处理。而2012年7月,N2处理群落高度最高(23.9 cm),N1次之(20.9 cm),且所有施肥处理均显著(P<0.05)高于N0;8月,N3处理群落高度最高(28.7 cm),N4处理次之(27.1 cm),分别比对照显著高出76.1%和66.3%(P<0.05),其它施肥处理也高于对照(P<0.05);在9月和10月,N4处理群落高度均为最高(29.5 cm和38.7 cm),分别比对照显著高出56.9%和100.5%(P<0.05),N0处理群落高度始终显著低于施肥处理(P<0.05)。
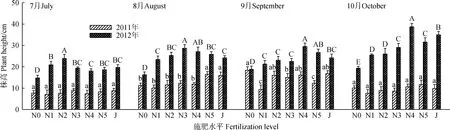
图2 施肥处理对轻度退化草地群落高度动态的影响Fig.2 Effects of fertilization on vegetation height of light degraded meadow
表3 不同处理高度均值Table 3 Mean plant height of different treatment/cm

年份Year月份Month7月July8月August9月September10月October20118.0b12.8b14.9b9.6b201219.3a24.4a23.7a29.3a
2.4 年份及施肥处理对轻度退化草地地上生物量的影响
年份及施肥处理对轻度退化草地地上生物量的影响见表1和表4。从年份上来看,2012年每月不同处理禾草类和杂草类生物量与2011年相比有高有低,而莎草类和总生物量则始终高于2011年;同时,2012年每月不同处理禾草类、莎草类和总生物量的均值均高于2011年,杂草类生物量则低于2011年。2012年7—10月,不同处理禾草类生物量的均值分别比2011年高出43.7%,67.9%,22.7%和128.2%,莎草类生物量的均值分别比2011年高出407.1%,145.0%,181.6%和184.2%,总生物量的均值分别比2011年高出93.7%,65.3%,74.1%和130.2%,杂草类生物量的均值分别比2011年降低68.7%,64.5%,27.0%和40.7%。可以看出,禾草类和杂草类对氮肥的响应较快,施肥后生物量迅速增加;莎草类生物量在年份间的差异较大,对氮肥的响应较慢,施肥后次年生物量大幅增加。从年度总生物量来看,禾草类的地上生物量在2011年以J处理最高(506.9 g·m-2),显著高于除N4外其它各处理(P<0.05),较对照高出96.6%,N4处理仅次于J处理,而2012年则以N4处理生物量最高,显著高于其它各处理,较对照高出207.4%;莎草类的地上生物量在2011年以N5处理最高,显著高于其它各处理(P<0.05),较对照高出51.8%,2012年则以N4处理最高,较对照高出61.6%;杂草类的地上生物量在两年均以N5处理最高,显著高于其它各处理(P<0.05),分别较对照高出151.4%和80.8%;年度总生物量在两年均以N4处理最高,显著高于其它各处理(P<0.05),分别较对照高出54.3%和100.8%。整体来看2012年年度总生物量高于2011年,其中N4处理2012年地上生物量是2011年的2.3倍,N0处理2012年地上生物量是2011年的1.8倍。
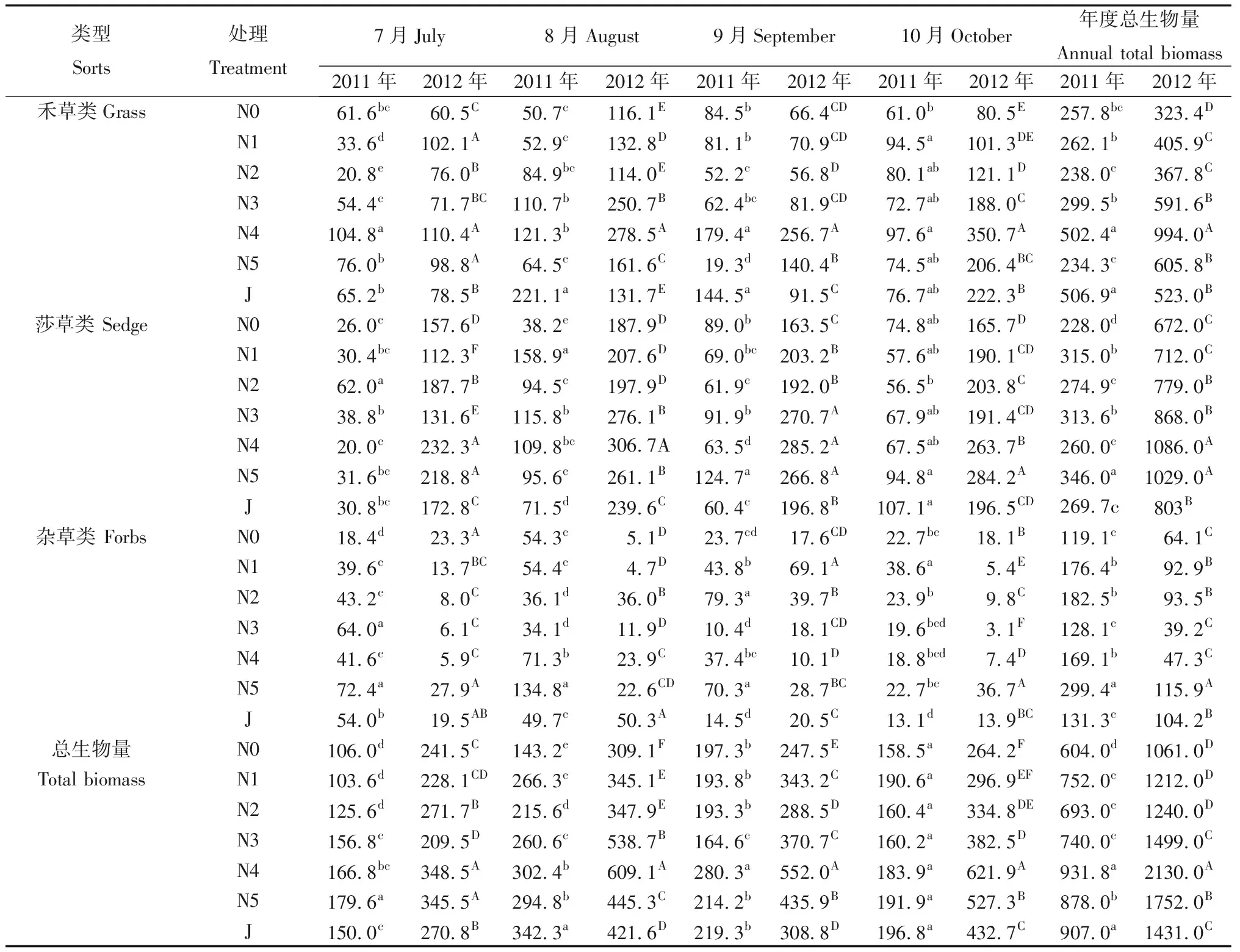
表4 施肥处理对轻度退化草地地上生物量的影响Table 4 Effects of fertilization treatments on over ground biomass of light degraded meadow/g·m-2
注:不同小写字母表示2011年同一月份不同施肥处理间差异显著(P<0.05),不同大写字母表示2012年同一月份不同施肥处理间差异显著(P<0.05),下同
Note:Different lowercase letters for the same month indicate significant difference among different years and treatments at 0.05 level in 2011,different uppercase letters for the same month indicate significant difference among different years and treatments at 0.05 level the in 2012
从不同施肥处理来看,随着牧草生育期的延长,2011年各经济类群和总生物量总体呈现先增后降的规律,最大生物量集中在8月;2012年则不同,莎草类和杂草类生物量总体呈现出先增后降的规律,而禾草类和总生物量总体呈现出先增后降再增的规律,不同处理的最大生物量在数量和出现时间上差异较大。2011年7月,总生物量N5处理为最高(179.6 g·m-2),其次为N4处理(166.8 g·m-2),但两者总生物量构成不同,N5处理以杂草类和禾草类为主,杂草类生物量显著(P<0.05)高于除N3外其它各处理,N4处理总生物量以禾草类为主,禾草生物量显著(P<0.05)高于各施肥处理。8月,J处理的禾草类生物量(221.1 g·m-2)和总生物量最高(342.3 g·m-2),显著(P<0.05)高于其它各处理,其次为N4处理(禾草生物量121.3 g·m-2和总生物量302.4 g·m-2);N5处理杂草类生物量(134.8 g·m-2)显著(P<0.05)高于其它各处理。9月时,N4处理禾草类生物量(179.4 g·m-2)和总生物量(280.3 g·m-2)均显著(P<0.05)高于其它各处理;10月时,各处理总生物量较9月均下降,N4处理禾草类生物量仍为最高,显著(P<0.05)高于对照,但与各施肥处理无差异(P>0.05);J处理莎草生物量和总生物量最高(107.1和196.8 g·m-2),较对照分别增加43.2%和24.2%。2012年7月,N4处理的总生物量最高(348.5 g·m-2),比对照高出44.3%,N5次之(345.5 g·m-2),显著(P<0.05)高于其它处理,两者的总生物量构成均以莎草类为主,分别占总生物量的66.7%和63.3%,同时,N4处理的杂草类生物量显著(P<0.05)低于N5,仅有5.9 g·m-2。8月时N4处理的禾草类、莎草类和总生物量均为最高(278.5,306.7和609.1 g·m-2),显著(P<0.05)高于其它各处理,比对照分别高出139.9%,38.7%和97.1%,杂草类生物量最低。9月仍以N4处理禾草类、莎草类和总生物量为最高(256.7,285.2和552.0 g·m-2),较对照高出286.6%,74.4%和123.0%,禾草生物量和总生物量显著(P<0.05)高于其它处理。10月时,N4、N5和J的总生物量达到最大值,且N4处理禾草生物量(350.7 g·m-2)和总生物量(621.9 g·m-2)均显著(P<0.05)高于其它各处理,比对照高出335.7%和135.4%,杂草类生物量显著(P< 0.05)低于对照。
3 讨论
植被盖度的降低、物种多样性的减少和群落组成成分的简单化是草地植被退化的主要特征,预示着生态环境的改变对植被的生存选择愈加严格[8]。草地植被退化使高寒草地生态系统结构与功能严重受损[20],研究草地植被退化机理及恢复措施有助于推动当地畜牧业的持续健康发展[21-22]。施肥和围栏封育是退化草地恢复的有效措施,在我国草地管理中得到广泛应用[23-24]。施肥可以提高青藏高原高寒草甸植物生产力,对草地恢复、草地生态系统稳定性具有促进作用[25],这在本研究中也得到很好地体现,与对照相比,施肥处理显著增加草地群落盖度、高度和生物量,但结果同时表明不同肥料和施用量的作用差异较大,不同经济类群对施肥的响应也有差异。
在退化草地施氮量方面已有多位学者进行研究,适量的氮肥增施量对于提高草地生产力具有积极意义,但过量增施氮肥不仅增加生产成本,而且易造成杂草旺长、优势种变换的现象从而影响畜牧生产,还会导致土壤板结等质量变差现象;最佳施氮量因地区而异,不同地区在退化草地管理中应结合生态和经济效益综合考虑,以达到增产增收的效果。陈亚明等[26]从维持草地营养品质考虑,认为甘南州高寒草地施肥量不应超过600 kg·hm-2水平,当施肥水平达到600 kg·hm-2时,豆科在群落地上生物量的比例比不施肥下降80.9%。曹文侠等[27]对天祝县轻度退化高寒草地的研究表明,施氮显著增加各功能群植物的高度和禾本科植物盖度,同时施氮可显著增加禾本科、莎草科、豆科地上生物量和总地上生物量,降低杂类草盖度和生物量,施肥量为308.6~385.8 kg·hm-2时效果最佳。景美玲等[28]对祁连县高寒草甸轻度退化草地的研究表明,在高寒草甸退化草地上添加氮素可显著提高植被盖度和地上生物量,添加150 kg·hm-2的尿素地上生物量2年内平均提高43.6%,投入产出最高,且对土壤养分有改善作用。本研究中,195 kg·hm-2(N4)施氮水平下退化草地的盖度、高度和地上生物量均为最佳,为本试验地区的适宜施肥浓度。当氮肥水平继续增加到240 kg·hm-2(N5)时,各经济类群和总地上生物量较N4下降,杂草生物量上升,说明高氮添加对草地的恢复不利。过量施肥将使土壤与植物营养元素失衡,造成土壤酸化,对植物产生毒害作用,降低群落生产力[29]。
在本研究中,在施肥当年和施肥第2年氮肥草地植被的高度和盖度均提高。氮肥是一种速效养分,极易在短时间内被具有较高氮素利用率的禾本科牧草吸收利用[30],地上部分迅速生长,高度和盖度增加,迅速占领上层空间;沈景林等[31]研究表明,在高寒退化草地施氮肥可使草群高度较对照增加26.8%~108.2%;在大通河上游祁连县退化草地开展的研究表明施肥处理草地群落高度较对照增加36.9%~202.8%,并推测可能由于该退化草地土壤对氮素供应的天然不足成为抑制植被生长的重要因素。施肥主要是增加了土壤速效养分,改变了植物竞争力,进而提高牧草产量,施氮肥消除了青藏高原氮素的限制作用,氮利用率高的禾草迅速生长,遮蔽了其他矮小物种,从而具有较强的获取地上光的能力[28]。施氮增加了土壤中可利用氮含量,增强了植物对碳的同化,从而加速了植物生长。施肥增加了土壤中的有效资源水平,从而导致群落生产力增加[25]。施肥使植物个体增高增壮,使得植被盖度整体上增加;陈子萱[32]研究表明氮肥、磷肥和有机肥混合施肥草地植被盖度可增加7%,其中氮肥对盖度的增加作用最大。还有研究表明施肥使禾草类个体数、盖度和高度增加,禾草竞争力增强,对其它种群造成抑制,杂类草减少,草地物种数减少、物种丰富度显著下降,Simpson指数和Shannon-Wiener指数减小,整体种间竞争降低。本研究中施氮处理的草地植被高度和盖度均增加的原因可能是施氮增加了土壤可利用氮的含量,氮素不再成为牧草生长的限制作用因子;退化草地的特征之一是草地土壤的有机质含量低,施肥还可能通过促进土壤中微生物的活动、调节土壤酶的活力、碳氮耦合等方式增加土壤有机质含量;施氮促进了牧草地上部分的生长,光合面积增大,光合产物合成旺盛使得其高度、盖度增加;施氮导致的毒杂草的数量和种类的减少解除了毒杂草对牧草的化感效应,减弱了种间竞争,促进牧草生长,高度和盖度增加。
微生物菌肥较化肥有对土壤的改良效果好、环保、利用率高等优点,可以疏松土壤,打破板结,有益微生物可能成为土壤环境中的优势菌,抑制病原菌和腐败菌的生长,避免土传病害的发生。除在大宗作物和蔬菜上开展了相关的研究外,也有对退化草地等方面的研究。卢建男等[33]发现嗜盐碱微生物菌肥施用量7 500 kg·hm-2,并结合底层衬膜技术改良兰州新区秦王川的盐碱土效果最佳,使该地区土壤适宜植物生长。柴青等[34]发现施用微生物菌肥可促进针茅(Stipacapillata)、扁穗冰草(Agropyroncristatum)和紫花苜蓿的生长,且对2种禾本科草种的影响更大。本研究中采用的EM菌由光合菌类、乳酸菌类、酵母菌类等三大菌群的多属多种微生物组成。通过发酵工艺将上述好氧及厌氧微生物混合培养,各微生物在其生长过程中产生有用物质及其分泌物,形成相互生长的基质和原料,通过相互共生、增殖关系形成一个结构稳定、功能广泛的具有多种多样微生物群落的生物菌群。EM菌中乳酸菌群能加速土壤中的木质素和纤维素成分的分解,而酵母菌群可以进一步利用这些分解产物生产出促进细胞分裂的活性化物质,为其他有效微生物增殖提供营养,在这两种菌群的共同作用下,EM菌群在土壤中表现良好的活力,从而产生了高的微生物生物量碳[35]。EM菌和病原微生物争夺营养的竞争,由于EM菌在土壤中极易生存繁殖,所以能较快而稳定地占据土壤中的生态地位,形成有益的微生物菌的优势群落,从而控制病原微生物的繁殖和对作物的侵袭。菌肥是生态农业的发展方向,更有利于农业的可持续发展。本研究发现菌肥对轻度退化草地盖度、高度和地上生物量均有促进作用,且在施肥后当年的效果整体优于氮肥处理,有起效快的优点,但成本较高,具体效益还有待进一步评估。
作物在不同年际间对肥料的利用效率方面有较多研究,特别是在控释肥一次性施入条件下作物的肥料利用效率[36]、转运、生长发育状况、土壤性质的变化方面[37-38]。张伟纳等[39]研究表明与农户习惯施肥(尿素一基一追)相比,一次性基施控释尿素增加作物产量进而起到控量节本、增产增效及提高氮素利用率的作用,可作为潮土区黄淮平原较适宜的集约化农田氮肥运筹方式。孙志考等[40]在大田试验条件下,研究了在冬小麦-夏玉米复种连作体系下氮肥周年运筹对其产量以及关键构成要素的影响,研究表明冬小麦-夏玉米复种连作体系下,中高氮肥水平对冬小麦产量关键性构成要素穗粒数、穗粒质量和每穗结实小穗数的影响均较对照和低氮肥水平有所提高,千粒质量下降;对夏玉米百粒质量、果穗粗、果穗长的影响具有显著提高,但中高氮肥之间差异不显著。在本研究中,施肥第2年轻度退化草地的植被高度、盖度和地上生物量均比第1年高,特别是每年的10月时差异最为明显。根据各类群草的地上生物量来看,莎草和禾草的增加量较高,莎草的地上生物量在所有处理下均表现为2012年高于2011年,而杂草类普遍发生地上生物量降低的现象,N2和N4处理7—10月均表现为2011年高于2012年,降低的幅度小于莎草科和禾本科草的增加幅度,所以在总生物量上表现为2012年高于2011年。造成这种年际间差异的原因,首先可能是种子库的影响,土壤种子库(是植物种群生活史的一个重要阶段,是指土壤上层枯落物和土壤中全部有活力种子的总和。它是植物群落更新发展的重要基础之一,为植物群落的演替、群落遭受干扰和破坏后的恢复提供繁殖体,在重塑和维持生物多样性方面具有重要的作用。采用土壤种子库研究植被退化恢复过程是种较为成功的方法[41]。2012年时上年施入的肥料可以为土壤种子库的有活力的种子提供更好的营养物质条件,可供更多的种子发芽,群落结构和草的生长状况更加良好,但2011年在6月施肥对于当年的草地群落结构而言影响较2012年。其次曹文侠等[27]指出施氮增加了土壤中可利用氮含量,增强了植物对碳的同化,从而加速了植物生长,进而有更多的凋落物分解后形成有机质进入土壤。在本研究中,2012年的植被受2011年枯落物的影响,2011年施肥处理的地上生物量较不施肥高,说明施肥对于地上生物量的增加发挥积极作用,在这种条件下,2011年枯落物中的可降解的氮素的量也较多,这也为2012年草的生长提供了营养条件。第三是氮肥运筹与肥料的持效性对土壤的影响,在2011年施入的氮肥部分在当年被草类吸收,部分暂时贮存于土壤中在2012年时被利用,在肥效缓慢释放的条件下,2012年草的整体生长状况较优,还有两年间种群结构发生差异,具有高生物量的种类增多,从而影响整体的生物量和植被特征。菌肥与高氮水平N4,N5相比表现出地上生物量略低的现象,可能与经过堆肥发酵的菌肥较化肥有更好的速效性有关。
在不同经济类群对施肥的响应上,本研究中,禾草对氮肥的响应较快,施肥后当年生物量迅速增加;莎草类对氮肥的响应较慢,施肥后次年生物量大幅增加;杂草生物量在初期上升明显,但后期又被抑制,这与已有的研究结果类似[42-43]。氮肥的施用消除了退化草地生态系统的养分限制,使得养分利用率高的禾本科植物迅速生长,高度和盖度增加,改变了群落结构和竞争格局,使植物之间的竞争从地下的养分转为地上空间和光竞争[44],限制了莎草和杂草的生长。宗宁等[45]比较了围栏内外施肥对退化草甸的影响,发现不管内外莎草生物量均增加,而禾草地上生物量在围栏内显著上升。到了次年,物种对施肥的响应与植物的物候期有显著相关性,6月试验区草地开始返青,莎草植物的物候期相对较早,能在其他植物密闭之前提早利用有效资源,以满足其生长发育的要求[42,46],因而次年莎草的生物量要高于禾草,且繁茂的莎草进一步抑制了杂草的生长。
4 结论
在贵南县轻度退化草地,施氮配合围栏封育能有效提高植被群落高度、盖度和地上生物量,改善植被结构组成,且次年效果更明显。195 kg·hm-2施氮量能显著提高次年整个牧草生长季草地植被群落盖度和高度,增加禾草类、莎草类地上生物量和总地上生物量。因此,195 kg·hm-2尿素施用量配合围栏封育的措施可用于贵南县轻度退化草地的植被恢复。
猜你喜欢
疯狂英语·新读写(2022年1期)2022-11-23
疯狂英语·读写版(2022年1期)2022-01-29
文史博览·文史(2021年1期)2021-04-01
黑龙江工程学院学报(2020年5期)2020-10-21
水土保持研究(2019年6期)2019-10-19
畜牧兽医科学(2019年24期)2019-09-10
南方农业·下旬(2017年11期)2018-01-09
绿色科技(2017年15期)2017-09-01
中国饲料(2017年15期)2017-08-22
杂草学报(2015年2期)2016-01-04
