
亚硝酸盐和氨对日本沼虾肝胰腺代谢的影响
2019-09-17 11:07余苗苗江敏吴昊吴丹金若晨孙世玉姚丹王旭娜
生态毒理学报 2019年3期
余苗苗,江敏,2,*,吴昊,吴丹,金若晨,孙世玉,姚丹,王旭娜
1. 上海海洋大学海洋生态与环境学院,上海 201306 2. 上海市水产养殖工程技术研究中心,上海 201306 3. 上海海洋大学水产与生命学院,上海 201306
日本沼虾(Macrobrachiumnipponensis)又名青虾、河虾,隶属甲壳纲类,十足目,长臂虾科,沼虾属[1],具有较高的食用价值和经济价值,是我国重要的淡水养殖经济虾类之一。据2017年《中国渔业统计年鉴》统计,我国虾类养殖产量203.21万t,其中日本沼虾27.26万t,较2016年增长2.84%[2]。在大规模的集约化养殖模式下,养殖管理措施不当会造成池塘底部残饵及粪便的积累,引起水质恶化,致使虾体产生应激反应,摄食能力和自身免疫下降,严重时会危害虾的生长与存活[3-4]。
氨与亚硝酸盐是水产养殖中最常见的污染物,两者来自于水生生物的直接代谢和环境中有机物的分解转化。其中,非离子氨会破坏水生动物的排泄系统及渗透平衡,损伤组织结构,致使代谢紊乱等,严重时可致动物死亡[5-7]。有研究表明,水中过高的亚硝酸盐进入鱼虾体内后,会导致血淋巴pH降低,进而影响生物体内氧的运输,扰乱氮排泄及损坏器官等[8-9]。肝胰腺是甲壳纲动物脂质储存和加工的重要器官,是消化、吸收和代谢的主要场所[10],也是受氨及亚硝酸盐毒性损害的主要部位之一。冼健安等[11]研究发现,斑节对虾肝胰腺中氨氮的含量显著高于血淋巴、鳃和肌肉中的含量,且更易于积累其中。蒋琦辰等[12]研究发现,随着氨氮浓度的升高,红螯光壳螯虾幼体肝胰腺中的可溶性蛋白和甘油磷脂会受到抑制,含量降低。Zhou等[13]证实氨氮胁迫会造成斑节对虾肝胰腺和鳃中几丁质(PmChi-4)含量的减少。虾体内相关酶的活性如超氧化物歧化酶(SOD)、酚氧化酶(PO)在亚硝酸氮的胁迫下会受到不同程度的影响,从而影响虾的生长代谢[14-15]。亚硝酸盐与氨对日本沼虾的胁迫会对其养殖产量产生较大影响[16-17],继而导致经济损失。近年来,关于亚硝酸盐和氨对于虾类的毒性研究,多集中在高浓度时对虾生长及生理活动、免疫功能等方面的影响。但在运用代谢组学探究低浓度亚硝酸盐和氨对日本沼虾的毒性机制方面的研究鲜有报道。本文基于养殖生产中通常浓度的亚硝酸盐、氨,从代谢组学角度探讨其对日本沼虾肝胰腺的影响,从而对指导虾类的养殖生产具有重要的现实意义。
1 材料与方法(Materials and methods)
1.1 实验材料
实验所用日本沼虾取自上海青浦区某养殖场,平均体质量(1.8±0.3) g,体长(0.5±0.2) cm。正式实验前,于水箱(长×宽×高=1.5 m×1.8 m×1.2 m,内含350 L经充分曝气除氯的自来水)中暂养一周。暂养期间控制水温为(21±0.5) ℃,pH为(8.2±0.1),溶解氧为(6.0±0.5) mg·L-1,每天定时投喂适量饵料,喂食30 min后更换1/4体积水,及时清除残饵及排泄物,暂养期间死亡率低于10%。
试剂:HPLC级甲醇(Sigma),HPLC级水(Sigma-Aldrich公司),N,O-双(三甲基硅烷基)乙酰胺(BSTFA),含1%茶渣微晶纤维素(TMCC)(上海安谱),分析纯吡啶(Sigma-Aldrich公司),分析纯甲氧基胺盐酸盐(Sigma-Aldrich公司),分析纯2-氯苯丙氨酸(Sigma-Aldrich公司)。分析纯NH4Cl和NaNO2(国药集团),用蒸馏水将NaNO2和NH4Cl配制成10 g·L-1的母液,现配现用。
仪器:液质联用仪(Waters, Q-Tof),气质联用仪(Algilent, 7890A/5975);全自动快速研磨仪(上海净信科技,Tissuelyser-24),离心机(Eppendor, Centrifuge 5810 R),高速离心浓缩仪(Labogene, 2236R),恒温混匀仪(杭州瑞诚仪器有限公司,TS100)。
1.2 实验方法
1.2.1 预实验
课题组对上海青浦、奉贤等区的淡水虾类池塘水质进行了持续监测,在所获近3万个数据中,一个养殖周期内虾塘氨氮通常在0.1~2.0 mg·L-1范围内,且多在1.0 mg·L-1附近波动;亚硝氮多为0.001~0.6 mg·L-1,且养殖初期即会快速升至0.25 mg·L-1以上。因此设置了3组不同浓度的亚硝酸盐,分别为对照组(≤0.05 mg·L-1)、低胁迫组(0.25 mg·L-1)、高胁迫组(0.5 mg·L-1);氨胁迫预实验共设3组,分别为对照组(pH=8.0,总氨氮(TAN)≤0.1 mg·L-1,即非离子氨氮小于0.005 mg·L-1)、低胁迫组(pH=8.6,TAN=1.0 mg·L-1,即非离子氨氮为0.152 mg·L-1)、高胁迫组(pH=9.0,TAN=1.0 mg·L-1,即非离子氨氮为0.378 mg·L-1)。每组实验水箱(体积48 cm×36 cm×28 cm)含有30 L经充分曝气处理的自来水,分别放入6尾日本沼虾,其余养殖管理同暂养期间,胁迫实验进行5 d后采集虾样。
取样时,迅速将日本沼虾置于液氮中快速冷冻,在冰上解剖取其肝胰腺组织,每尾虾取其肝胰腺样本约30 mg于离心管中作为一个实验样本,在装有样品的离心管内加入20 μL内标化合物(0.3 mg·L-1的2-氯苯丙氨酸)以及800 μL提取液(甲醇∶水=4∶1,V/V),匀浆后于-20 ℃冰箱静置10 min后取出,10 000 r·min-1离心10 min,取200 μL上清液于色谱瓶中用于液质联用的检测;另取200 μL上清液于样品瓶中,干燥后经BSTFA硅烷化,用于气质联用检测。
1.2.2 正式实验
对预实验结果进行主成分分析,与对照组相比,对日本沼虾肝胰腺代谢产物有显著影响的实验组为亚硝酸盐氮浓度为0.5 mg·L-1和高浓度氨(pH=9.0,TAN=1.0 mg·L-1,即非离子氨0.378 mg·L-1)胁迫组。因此,后续亚硝酸盐胁迫设4个不同胁迫时间组:对照组(C)、胁迫2 d实验组(NO2-2d)、胁迫3 d实验组(NO2-3d)与胁迫5 d实验组(NO2-5d),其中亚硝酸盐氮浓度均为0.5 mg·L-1;pH=9.0,TAN为1.0 mg·L-1条件下设4个不同胁迫时间组:对照组(C)、胁迫2 d实验组(TAN-2d)、胁迫3 d实验组(TAN-3d)和胁迫5 d实验组(TAN-5d),各组设定胁迫时间截止时采集虾样,样品处理方法同“1.2.1”。
1.2.3 定量实验
经过对pH=9.0,TAN=1.0 mg·L-1的氨胁迫2 d实验组(TAN-2d)差异代谢物的鉴定及代谢通路分析,在与对照组有显著性差异的化合物中发现了草酰乙酸、α-酮戊二酸等三羧酸循环的相关代谢物。因此,对pH=9.0、TAN为1.0 mg·L-1的氨氮组进行进一步实验,设置胁迫时间分别为12 h、24 h和48 h,采用气质联用仪对日本沼虾肝胰腺中的草酰乙酸、柠檬酸、L-谷氨酸、α-酮戊二酸进行定量测定。截止时间采集虾样,样品处理方法同“1.2.1”。
1.3 气相、液相色谱-质谱联用仪条件
气相色谱-质谱联用仪(Algilent, 7890A/5975)检测使用HP-5MS毛细管柱(60 m×0.25 mm×0.25 μm);进样体积为1 μL,进样口温度为260 ℃,初始温度为60 ℃,以10 ℃min-1的速度升到315 ℃,恒定流速1.0 mL·min-1;EI离子源能量70 eV,离子源温度230 ℃,四极杆温度150 ℃,质量扫描范围70~600 amu;以全扫描模式获得总离子流色谱图。
液质联用(Waters, Q-Tof),流动相0.1%甲酸水溶液(A)-乙腈(B),梯度洗脱条件:0~2 min,2% B;2~10 min,100% B;10~12 min,100% B;12.1 min,2% B;12.1~15 min,2% B。流速0.3 mL·min-1,进样体积2 μL。
1.4 数据分析
气质联用及液质联用数据由Mass Hunter软件和QI软件进行归一化处理,并获得样品代谢组份及丰度的矩阵信息。用SIMCA 14.1软件,对获得的数据进行PCA和OPLS-DA分析并筛选得到差异代谢物,在OPLS-DA分析结果的S-Splot图中,选择∣p(corr)∣>0.5、∣p∣>0.1、VIP值>1.0且P值<0.05的差异代谢物。通过NIST谱库检索,利用HMDB、Lipids和KEGG等在线数据库对差异代谢物的结构、名称进行比对确认,首先结合气相色谱-质谱联用仪、液质联仪用所得结果,筛选出实验组中与对照组有显著差异性的代谢物,再将所鉴定出的带有HMDB和KEGG编号的差异性代谢物输入Metabo Analyst 3.0在线通路分析数据库,并依次查看差异性代谢物在KEGG数据库中所对应的代谢通路。
2 结果(Results)
2.1 亚硝酸盐对日本沼虾肝胰腺代谢的影响
经预实验后选择了亚硝酸盐氮浓度为0.5 mg·L-1来进行胁迫实验,时间设置分别为2、3和5 d。检测结果经主成分分析(PCA)(图1),对照组与实验组(NO2-2d)间的代谢轮廓呈明显的分离趋势,即经0.5 mg·L-1亚硝酸盐胁迫2 d后,日本沼虾肝胰腺代谢和对照组之间产生了显著差异;而胁迫3 d与5 d组则与对照组区分不明显,推测胁迫3 d、5 d后,其新陈代谢有恢复的可能。随后,对对照组(C-2d)和实验组(NO2-2d)进行重复性验证,用正交偏最小二乘法-判别分析(OPLS-DA)对肝胰腺的代谢数据作进一步分析,结果见图2和图3,表示模型解释能力的参数RX2=0.834,表示模型预测能力的参数Q2=0.712,说明模型可靠。由此可知模型对自变量的拟合能力和预测能力良好,对照组与实验组在肝胰腺代谢谱中均可以明显区分。
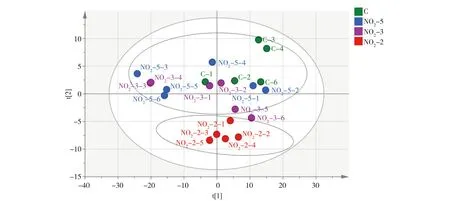
图1 日本沼虾经0.5 mg·L-1的亚硝酸盐胁迫后肝胰腺的代谢化合物主成分(PCA)得分图注:C代表对照组,NO2-5代表亚硝酸盐(0.5 mg·L-1)胁迫5 d的实验组,NO2-3代表亚硝酸盐(0.5 mg·L-1)胁迫3 d的实验组, NO2-2代表亚硝酸盐(0.5 mg·L-1)胁迫2 d的实验组,NO2-5-x、NO2-3-x和NO2-2-x代表各组内的平行样本,即第1至第6尾虾。Fig. 1 PCA score plots of metabolites in Macrobrachium nipponensis hepatopancreas in control group and experimental groups exposed to NO2-N (0.5 mg·L-1) Note: C is control group; NO2-5 is experimental group exposed to NO2-N (0.5 mg·L-1) for 5 d; NO2-3 is experimental group exposed to NO2-N (0.5 mg·L-1) for 3 d; NO2-2 is experimental group exposed to NO2-N (0.5 mg·L-1) for 2 d; NO2-5-x, NO2-3-x and NO2-2-x is a parallel sample of each group, from the frist shrimp to the sixth.
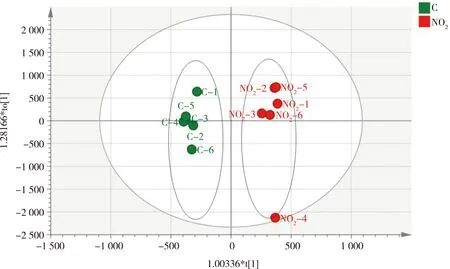
图2 日本沼虾经0.5 mg·L-1亚硝酸盐胁迫2 d后,实验组与对照组肝胰腺代谢化合物的正交偏最小二乘法(OPLS-DA)得分图注:C代表对照组,NO2代表亚硝酸盐(0.5 mg·L-1)胁迫2 d的实验组,NO2-x是实验组内的平行样本,即第1至第6尾虾。Fig. 2 OPLS-DA score plots of metabolites in Macrobrachium nipponensis hepatopancreas in control group and experimental groups exposed to NO2-N (0.5 mg·L-1) for 2 d Note: C is control group; NO2 is experimental group exposed to NO2-N (0.5 mg·L-1) for 2 d; NO2-x is a parallel sample of each group, from the frist shrimp to the sixth.
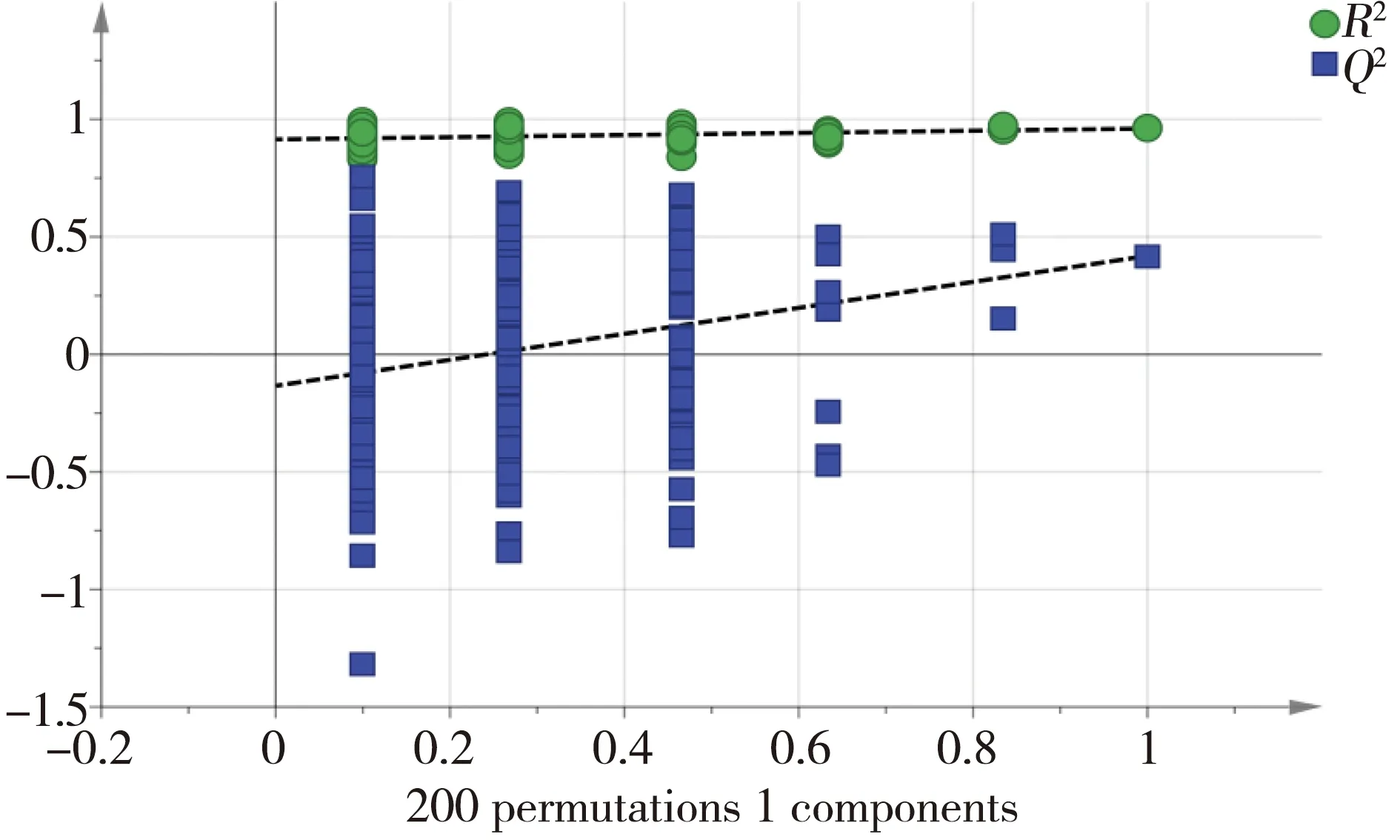
图3 OPLS-DA得分图(图2)的交互验证图注:R2表示累计方差值,Q2表示累计交叉有效性。Fig. 3 Permutation test of OPLS-DA score plots (figure 2) Note: R2 stands for cumulative variance; Q2 stands for cumulative cross validity.

表1 0.5 mg·L-1亚硝酸盐胁迫组(2 d)与对照组日本 沼虾肝胰腺代谢物中具有显著差异的成分列表Table 1 Differential metabolites in Macrobrachium nipponensis hepatopancreas between 0.5 mg·L-1 NO2-2 (2 d) exposure group and control group
注:↑ 表示相对于对照组含量升高,↓ 表示相对于对照组含量降低。
Notes: ↑ indicates increasing content compared with the control group; ↓ indicates decreasing content compared with the control group.
2.2 受亚硝酸盐胁迫的日本沼虾肝胰腺中差异代谢物的鉴定及代谢通路分析
经液质联用仪、气质联用仪分析检测,筛选出实验组与对照组间具有显著差异性的代谢物(表1),并进行在线通路分析,结果显示:这些差异性代谢物主要涉及氨基酸代谢、磷脂代谢、淀粉和蔗糖等的代谢;其中包含15条代谢通路,即苯丙氨酸、酪氨酸和色氨酸的生物合成(impact=1),缬氨酸、亮氨酸和异亮氨酸的生物合成(impact=0.666),甘氨酸、丝氨酸和苏氨酸代谢(impact=0.58),甘油磷脂代谢(impact=0.231),丙氨酸、天冬氨酸和谷氨酸代谢,谷胱甘肽代谢,苯丙氨酸代谢,组氨酸代谢,β-丙氨酸代谢,精氨酸和脯氨酸代谢,硫代谢,嘧啶代谢,嘌呤代谢,色氨酸代谢和酪氨酸代谢等。
2.3 氨对日本沼虾肝胰腺代谢的影响
经预实验后选择了pH=9.0,TAN=1.0 mg·L-1的氨氮组,进行了为期2、3和5 d的胁迫实验。检测结果经主成分分析(PCA)后表明,对照组与实验组TAN-2d 间的代谢物轮廓呈明显的分离趋势(图4),即对照组与氨氮(TAN-2d)组已产生显著差异。故随后选取(TAN-2d)组,对对照组和实验组(TAN-2d)进行重复性验证实验,用OPLS-DA法进一步分析肝胰腺的代谢数据(图5和图6),表示模型解释能力的参数RX2=0.814,表示模型预测能力的参数Q2=0.713,说明模型可靠。由此可知,模型对自变量的拟合能力和预测能力良好,对照组与实验组在肝胰腺代谢谱中均可以明显区分。
2.4 受氨胁迫的日本沼虾肝胰腺中差异代谢物的鉴定及代谢通路分析
经液质联用仪、气质联用仪分析检测,筛选出对照组与氨氮组间具有显著差异性的代谢物(表2)并进行在线通路分析。结果显示,这些差异性代谢物主要涉及氨基酸代谢、磷脂代谢、淀粉和蔗糖等代谢;其中包含17条代谢通路,即苯丙氨酸、酪氨酸和色氨酸的生物合成(impact=1),缬氨酸、亮氨酸和异亮氨酸的生物合成(impact=0.670),甘氨酸、丝氨酸和苏氨酸代谢(impact=0.580),甘油磷脂代谢(impact=0.231),柠檬酸循环(impact=0.182),缬氨酸、亮氨酸和异亮氨酸降解,半胱氨酸和蛋氨酸代谢,丙氨酸、天冬氨酸和谷氨酸代谢,甲烷代谢,谷胱甘肽代谢,苯丙氨酸代谢,组氨酸代谢,精氨酸和脯氨酸代谢,硫代谢,色氨酸代谢,酪氨酸代谢和甘油磷脂代谢等。
2.5 氨胁迫对日本沼虾柠檬酸循环的影响
在亚硝酸盐与氨胁迫下,日本沼虾肝胰腺中差异代谢物主要涉及氨基酸代谢、脂肪酸代谢、磷脂代谢和糖酵解等;在氨氮组与对照组有显著性差异的化合物中发现了草酰乙酸、α-酮戊二酸等三羧酸循环的相关代谢物。氨中毒会导致三羧酸循环的中断,阻碍能量供应,造成水生生物的昏迷或死亡等[18]。因此,设计了pH=9.0、TAN为1.0 mg·L-1,胁迫时间分别为12、24和48 h的胁迫实验,采用气质联用仪对日本沼虾肝胰腺中的草酰乙酸、柠檬酸、L-谷氨酸和α-酮戊二酸进行定量测定。
草酰乙酸与乙酰-CoA缩合形成三羧酸循环通路中的草酰乙酸、柠檬酸含量见图7,谷氨酸生成α-酮戊二酸通路中的谷氨酸、α-酮戊二酸含量见图8。利用Graphpad prism7.0作图,其中T检验的P≤0.05表明具有显著差异性。胁迫12 h时,日本沼虾肝胰腺中草酰乙酸含量与对照组相比显著升高,胁迫进行24 h与48 h后其含量回落,与对照组相比无显著差异;柠檬酸和谷氨酸含量与对照组相比,在12、24和48 h时差异均不显著;α-酮戊二酸的含量在12 h时与对照组相比有一定的上升,胁迫进行24 h及48 h后回落。
3 讨论(Discussion)
亚硝酸盐和氨是虾类养殖中重要的环境污染因子,胁迫后可使甲壳类生物产生组织病变,其中对肝胰腺的影响最为严重[19]。研究表明,亚硝酸盐和氨氮长时间胁迫可使甲壳纲动物的免疫力下降,肝胰腺组织结构受到破坏[20]。钟君伟等[21]对克氏原螯虾进行胁迫实验,发现高浓度的亚硝酸盐致使肝胰腺中总超氧化物歧化酶(T-SOD)和过氧化氢酶(CAT)活性降低,酸性磷酸酶(ACP)和碱性磷酸酶(AKP)活性在12 h时被激活,之后恢复到对照水平。李永等[22]研究发现,低浓度氨氮胁迫下,斑节对虾在实验前期生物免疫力增高,中后期机体生理失调,酶活力下降。本实验中,亚硝酸盐及氨胁迫组与对照组相比,日本沼虾肝胰腺中色氨酸含量升高,苯丙氨酸、酪氨酸等芳香族氨基酸的含量均降低,亚硝酸盐和氨可能造成了苯丙氨酸、酪氨酸和色氨酸合成与代谢的紊乱。亮氨酸是支链氨基酸(BCAA)中唯一的生酮氨基酸,缬氨酸属于生糖氨基酸,可共同保护肌肉,对血红蛋白合成及血糖水平调节有重要作用[23-25],且具有促进组织损伤修复等功能[26]。与对照组相比,亚硝酸盐胁迫组与氨胁迫组亮氨酸的含量均降低,表明亚硝酸盐与氨胁迫可能会抑制日本沼虾肝胰腺血红蛋白的合成及血糖调节。
通过Metabo Analyst 3.0通路分析,日本沼虾肝胰腺中的甘油磷脂代谢涉及磷脂酸(PA)、磷脂酰胆碱(PC)和磷脂酰乙醇胺(PE)。其中磷脂酸具有活化细胞、维持新陈代谢、增强免疫力等功能,磷脂酰胆碱和磷脂酰乙醇胺是构成细胞膜的主要成分,并且三者之间可相互转化[27]。亚硝酸盐与氨氮胁迫均能够引起三者脂质代谢紊乱,一旦出现紊乱易引起各种疾病的发生。与对照组相比氨氮胁迫组中鞘磷脂含量降低。鞘磷脂构成的多层膜结构对神经纤维起保护和绝缘作用,可水解为神经酰胺,在细胞膜上形成神经酰胺的富集区,增加肝细胞对各种刺激的敏感性,影响肝细胞的增殖、存活及凋亡[28-29]。推测在氨氮胁迫下日本沼虾肝胰腺细胞的增殖、存活及凋亡会受到一定影响。二酰基甘油三甲基高丝氨酸(DGTS)为细胞膜无磷脂[30-31],与对照组相比,0.5 mg·L-1亚硝酸盐与氨氮(pH=9.0,TAN=1.0 mg·L-1)胁迫组日本沼虾肝胰腺中DGTS含量均有所升高,表明其可能对肝胰腺细胞膜的组成产生影响。亚硝酸盐胁迫组与对照组相比具有显著差异性代谢物还包括:甘油二酯(DG),属于甘油酯,是动物组织中的微量成分,在血淋巴系统和肝脏中可相互转化,对于细胞膜具有一定的破坏作用[32-33]。亚硝酸盐实验组中DG含量显著升高,推测亚硝酸盐可能会对日本沼虾肝胰腺细胞膜造成影响。

表2 氨(TAN=1.0 mg·L-1,pH=9.0)胁迫组(2 d)与 对照组日本沼虾肝胰腺代谢物中具有显著差异的成分列表Table 2 Differential metabolites in Macrobrachium nipponensis hepatopancreas between ammonia (TAN=1.0 mg·L-1, pH=9.0, 2 d) exposure group and control group
注:↑ 表示相对于对照组含量升高,↓ 表示相对于对照组含量降低。
Notes: ↑ indicates increasing content compared with the control group; ↓ indicates decreasing content compared with the control group.
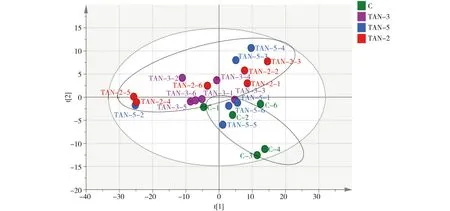
图4 日本沼虾经氨氮(TAN=1.0 mg·L-1,pH=9.0)胁迫后肝胰腺的代谢化合物主成分(PCA)得分图注:C代表对照组,TAN-5代表氨氮(TAN=1.0 mg·L-1,pH=9.0)胁迫5 d的实验组,TAN-3代表氨氮(TAN=1.0 mg·L-1,pH=9.0)胁迫3 d的实验组, TAN-2代表氨氮(TAN=1.0 mg·L-1,pH=9.0)胁迫2 d的实验组,TAN-5-x、TAN-3-x和TAN-2-x代表各组内的平行样本,即第1至第6尾虾。Fig. 4 PCA score plots of metabolites in Macrobrachium nipponensis hepatopancreas in control group and experimental groups exposed to ammonia (TAN=1.0 mg·L-1, pH=9.0) Note: C is control group; TAN-5 is experimental group exposed to ammonia (TAN=1.0 mg·L-1, pH=9.0) for 5 d; TAN-3 is experimental group exposed to ammonia (TAN=1.0 mg·L-1, pH=9.0) for 3 d; TAN-2 is experimental group exposed to ammonia (TAN=1.0 mg·L-1, pH=9.0) for 2 d; TAN-5-x, TAN-3-x and TAN-2-x is a parallel sample of each group, from the frist shrimp to the sixth.
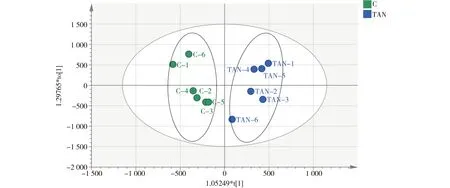
图5 日本沼虾经氨氮(TAN=1.0 mg·L-1,pH=9.0)胁迫2 d后,实验组与对照组肝胰腺代谢 化合物的正交偏最小二乘法(OPLS-DA)得分图注:C代表对照组,TAN代表氨氮(TAN=1.0 mg·L-1,pH=9.0)胁迫2 d的实验组,TAN-x代表实验组内的平行样本,即第1至第6尾虾。Fig. 5 OPLS-DA score plots of metabolites in Macrobrachium nipponensis hepatopancreas in control group and experimental groups exposed to ammonia (TAN=1.0 mg·L-1, pH=9.0) for 2 d Note: C is control group; TAN is experimental group exposed to ammonia (TAN=1.0 mg·L-1, pH=9.0) for 2 d; TAN-x is a parallel sample of each group, from the frist shrimp to the sixth.
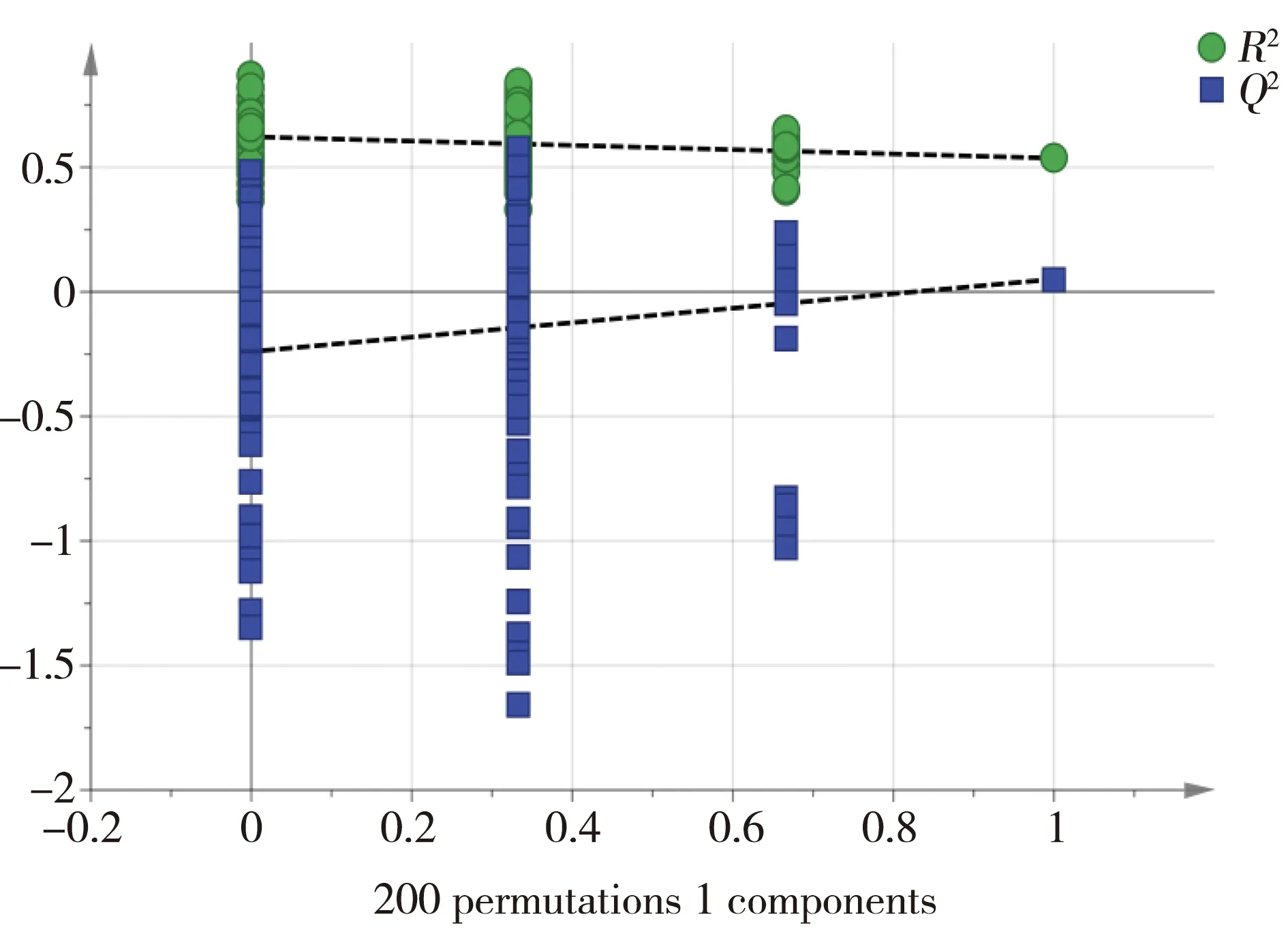
图6 OPLS-DA得分图(图5)的交互验证图注:R2表示累计方差值,Q2表示累计交叉有效性。Fig. 6 Permutation test of OPLS-DA score plots (figure 5) Note:R2 stands for cumulative variance; Q2 stands for cumulative cross validity.
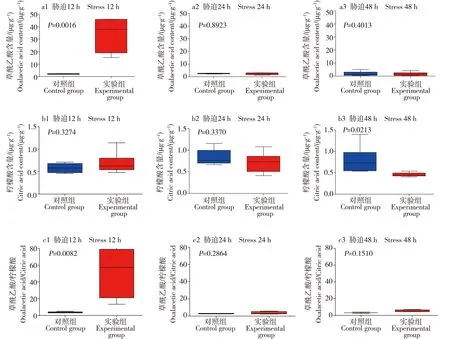
图7 对照组与氨胁迫组(pH=9.0,TAN=1.0 mg·L-1)12、24和48 h时日本沼虾肝胰腺中草酰乙酸和柠檬酸含量的变化Fig. 7 The changes in the contents of oxalacetic acid and citric acid in Macrobrachium nipponensis hepatopancreas at 12, 24 and 48 h in control group and experimental group (pH=9.0, TAN=1.0 mg·L-1)
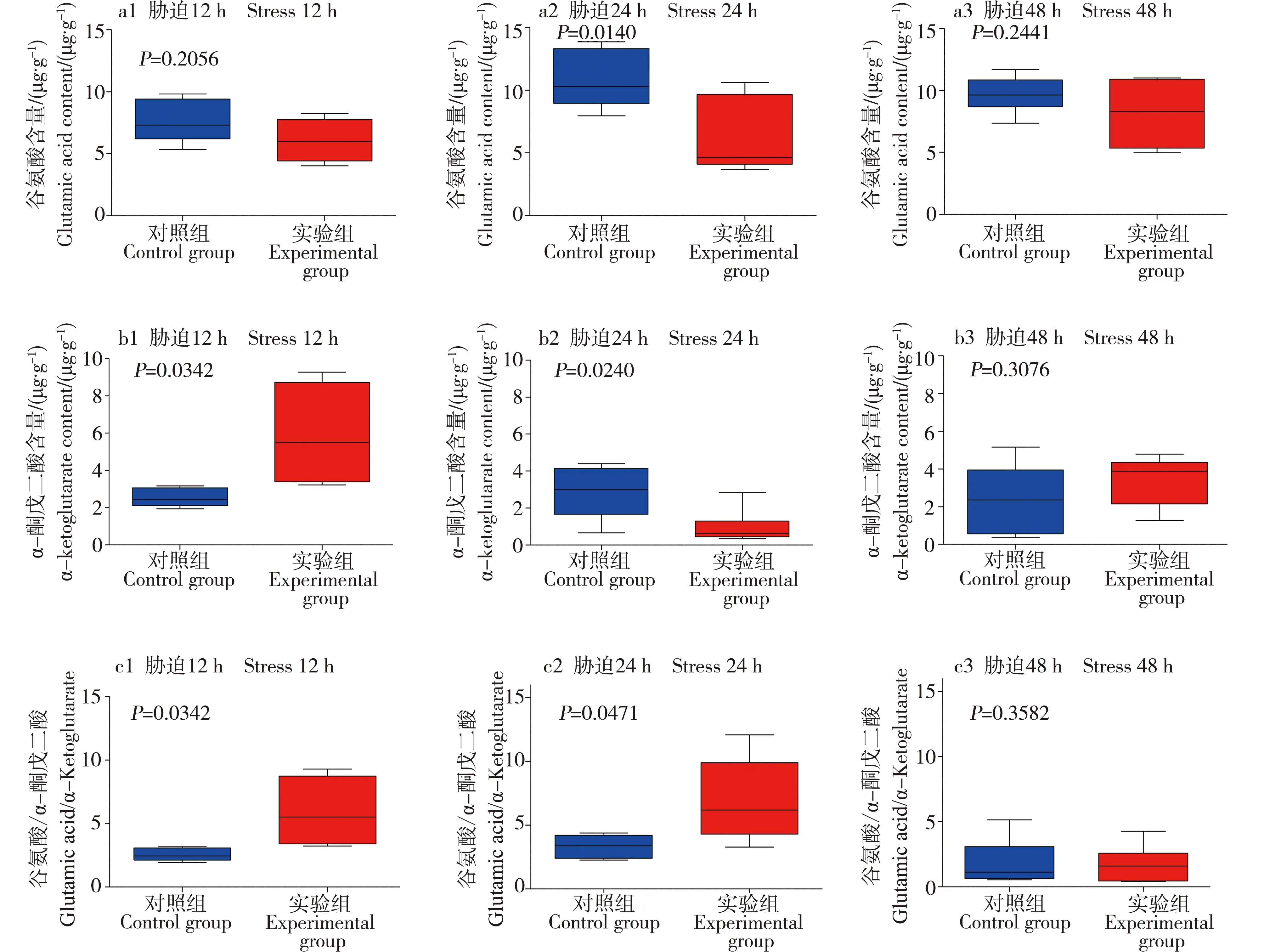
图8 对照组与氨胁迫组(pH=9.0,TAN=1.0 mg·L-1)12、24、48 h时日本沼虾肝胰腺中谷氨酸、α-酮戊二酸含量的变化Fig. 8 The changes in the contents of glutamic acid and α-ketoglutarate in Macrobrachium nipponensis hepatopancreas at 12, 24 and 48 h in control group and experimental group (pH=9.0, TAN=1.0 mg·L-1)
草酰乙酸与乙酰辅酶A在柠檬酸合酶(citrate synthase, CS)的催化作用下缩合形成柠檬酸,其中柠檬酸合酶属于调控酶,也是三羧酸循环中第一关键酶,是细胞内多种代谢途径的关键限速酶及代谢变化的标志物,CS具有高度的底物特异性,仅催化乙酰辅酶A与草酰乙酸缩合成柠檬酸,并且CS的活性强弱直接关系到柠檬酸的合成[34-37]。对pH=9.0,TAN=1.0 mg·L-1实验组进行进一步实验,对三羧酸循环中草酰乙酸、柠檬酸、谷氨酸和α-酮戊二酸进行定量测定,胁迫12 h的日本沼虾肝胰腺中草酰乙酸含量与对照组相比有显著上升,而产物中的柠檬酸含量变化不明显,可能氨胁迫使柠檬酸合酶的活性降低,抑制了产物中柠檬酸形成,从而使得作为底物的草酰乙酸含量得以积累,而合成的柠檬酸含量则未见增加。胁迫24 h和48 h时,日本沼虾体内的草酰乙酸含量与对照组差异不显著,推测在较长时间胁迫下虾体产生了适应性,并且有恢复的可能。α-酮戊二酸(α-KG)是一种重要的短链羟酸,是三羧酸循环重要的代谢中间产物[38],α-KG可通过转氨基形成谷氨酸从而连接细胞内碳-氮代谢,可直接参与集体的氧化供能及生物体内多种物质的化学合成[39-40]。在pH=9.0,TAN=1.0 mg·L-1条件下胁迫12、24和48 h后,日本沼虾肝胰腺内α-KG含量在胁迫12 h时有小幅度升高,可能在氨的胁迫作用下日本沼虾产生了短暂的应激反应。
综上所示:亚硝酸盐(0.5 mg·L-1)胁迫下,鉴定出49个与对照组相比具有显著差异的代谢化合物,亚硝酸盐会导致日本沼虾氨基酸代谢、脂肪酸代谢和甘油磷脂代谢紊乱。pH=9.0时,TAN(1.0 mg·L-1)胁迫后,鉴定出显著差异性代谢化合物52个,氨胁迫12 h会造成柠檬酸循环中的草酰乙酸与α-酮戊二酸的含量升高,氨会影响日本沼虾的三羧酸循环、氨基酸代谢、脂肪酸代谢和甘油磷脂代谢。后续可对氨氮胁迫下日本沼虾肝胰腺中的苹果酸脱氢酶及柠檬酸合酶的变化、以及磷脂代谢所受影响等作进一步验证,从代谢组学层面为氨对日本沼虾肝胰腺的毒性机制补充信息。
猜你喜欢
现代临床医学(2022年4期)2022-09-29
当代水产(2022年7期)2022-09-20
当代水产(2022年6期)2022-06-29
当代水产(2021年8期)2021-11-04
中国医院用药评价与分析(2020年3期)2020-05-29
农药科学与管理(2019年5期)2019-08-13
当代水产(2019年6期)2019-07-25
当代水产(2018年12期)2018-05-16
海洋与渔业(2017年11期)2018-01-26
恋爱婚姻家庭·青春(2016年7期)2016-07-05
