
柑橘木虱成虫繁殖行为前期及活动规律研究
2019-09-10 07:22黎海霖郑霞林王小云陆温
南方农业学报 2019年9期
黎海霖 郑霞林 王小云 陆温



摘要:【目的】明确柑橘木虱[Diaphorina citri(Kuwayama)]雌雄成虫交配前期和雌虫产卵前期的时长及成虫交配、产卵和羽化的昼夜节律,为探索更经济、有效和环保的柑橘木虱防控措施和性诱防控技术提供参考。【方法】采用活体九里香幼苗和成株为试验材料,以自制塑料杯及养虫笼为试验载具,在室内控制环境条件及室外自然狀态下对柑橘木虱成虫的繁殖行为进行观察,记录柑橘木虱成虫繁殖行为前期及繁殖行为昼夜节律数据。【结果】柑橘木虱雌雄成虫的交配前期分别为10.64±2.25和9.84±1.79 d;雌虫的产卵前期为12.58±2.90 d。成虫交配、产卵和羽化活动主要在2:00—21:00进行,其活动高峰期分别为9:00—10:00、11:00—12:00和9:00—10:00。【结论】柑橘木虱雌雄成虫交配、雌虫产卵和若虫羽化集中在白昼进行,雌雄成虫交配前期和成虫羽化日节律存在两性差异现象。可根据柑橘木虱成虫繁殖行为日节律的特点对其进行防治及预测预报。
关键词: 柑橘木虱;交配;产卵;羽化
中图分类号: S433.39 文献标志码: A 文章编号:2095-1191(2019)09-2009-06
Abstract:【Objective】To determine length of time of the pre-mating, pre-oviposition between male and female adults of Diaphorina citri(Kuwayama), and obtain the data of the mating and oviposition circadian daily rhythm of male and female adults, and data of eclosion circadian daily rhythm of nymph, so as to provide reference for economical,effective and green controlling of D. citri population as well as control technique by sex attractants. 【Method】Murraya jasminorage seedlings and adult plants were used as materials, and self-made plastic cups and insect cages were as carriers.The data of pre-reproductive behavior and the circadian rhythm of reproductive behavior of D. citri adults were obtained by observing the reproductive behavior of D. citri adults under indoor control environment condition and outdoor natural conditions. 【Result】The pre-mating periods of male and female adults were 10.64±2.25 and 9.84±1.79 d respectively, and the pre-oviposition of female was 12.58±2.90 d. Mating, oviposition and eclosion of adults were mainly carried out from 2:00 to 21:00, and their peak periods were 9:00 to 10:00, 11:00 to 12:00 and 9:00 to 10:00, respectively. 【Conclusion】The mating between male and female adults and oviposition of female adults and nymph eclosion are concentrated in the daytime, and the daily rhythm of pre-mating and eclosion is different between sexes. The control and forcast can be conducted based on the daily rhythm of D. citri reproductive behaviors.
Key words: Diaphorina citri(Kuwayama); mating; oviposition; eclosion
0 引言
【研究意义】柑橘木虱[Diaphorina citri(Kuwayama)]隶属于半翅目(Hemiptera)扁木虱科(Liviidae),是芸香科植物上的主要害虫,在全球柑橘种植产区均有分布(Stockton et al., 2017),在我国广泛分布于广西、广东、福建、海南、江西和台湾等地。柑橘木虱是柑橘黄龙病的传播媒介,常导致柑橘黄龙病暴发流行,对柑橘产业造成毁灭性打击(贾志成等,2015;Rwomushana et al., 2017);同时,因黄龙病毁灭的柑橘类果园在之后的10年将无法创造利润(Brar et al., 2015)。柑橘木虱成虫繁殖行为决定其种群的发生动态,包括成虫交配产卵日节律、成虫羽化日节律及种群年发生代数等,且成虫可通过取食传播柑橘黄龙病,因此,研究柑橘木虱成虫繁殖行为规律有助于进一步控制柑橘木虱种群数量,进而防控黄龙病的蔓延,对保障柑橘产业健康发展具有重要意义。【前人研究进展】近年来,有关柑橘木虱成虫繁殖行为的研究取得一定进展,特别是引入了分子生物学技术研究成虫的行为机制。Onagbola等(2008)使用电镜对柑橘木虱成虫的触角进行扫描观察,明确了柑橘木虱成虫触角上能进行远距离定位配偶的感器类型。Erikj等(2009)通过声波记录仪研究柑橘木虱雄成虫求偶过程所发出的声波信号,确定柑橘木虱雄成虫在较近距离主要利用声波对雌成虫进行配偶定位,且雌雄成虫存在多次交配现象。肖培彬等(2017)通过解剖不同虫龄亚洲柑橘木虱雌雄成虫的生殖器官,掌握了柑橘木虱成虫内生殖系统形态的变化规律。李超峰等(2019)通过解剖柑橘木虱成虫的复眼,明确了柑橘木虱成虫为日行性昆虫。Ibanez等(2019)研究了vga1和kr-h1基因在柑橘木虱雌虫产卵阶段的重要性,并解释了柑橘木虱不同个体繁殖能力存在差异的机制。Kelley等(2019)通过分子研究明确了黄龙病细菌可促进柑橘木虱扩散和繁殖。Qasim等(2019)通过分子研究来了解柑橘木虱在黄龙病植株上的发育和死亡率,明确了黄龙病细菌可支持柑橘木虱从植物韧皮部吸收更多的汁液。【本研究切入点】柑橘木虱成虫繁殖行为活动具有一定的日节律现象,在特定时间段防治成虫不仅能提高防治效果,还可减少农药的使用量。目前对柑橘木虱成虫繁殖行为特性及成虫繁殖行为活动日节律尚无详细文献记录。【拟解决的关键问题】采用活体九里香幼苗和成株为试验材料,以自制塑料杯及养虫笼为试验载具,通过室内和室外试验详细记录柑橘木虱成虫繁殖特性及繁殖活动规律,以了解柑橘木虱成虫需要经历交配和产卵前期的时长及成虫交配、产卵、羽化昼夜节律,为探索更经济、有效和环保的柑橘木虱防控措施和性诱防控技术研究提供参考。
1 材料与方法
1. 1 试验材料
1. 1. 1 供试虫源 从广西大学柑橘园的九里香植株上采集一定数量的柑橘木虱成虫,室外用5个养虫笼饲养,待雌虫交配产卵,若虫羽化为成虫即可获得试验虫源。取300头羽化后8~11 d的雌蟲和300头羽化后9~11 d的雄虫放入室外养虫笼内,用有嫩梢的活体九里香植株饲养雌雄成虫,经3 d饲养适应后作为繁殖行为活动昼夜节律观察试验虫源。
1. 1. 2 试验设备与环境条件 用圆柱形塑料杯作为观察成虫交配和雌虫产卵的载具,其由一个高9.50 cm、宽21.00 cm圆柱形塑料底座,一个高14.00 cm、杯口宽9.50 cm、底部宽5.50 cm的圆柱形塑料杯和1支50 mL指形管组成(图1)。观察设备为佳能6D相机和尼康SMZ445体视镜。室内试验环境控制在温度(25±1)℃、相对湿度(75±5)%、光照∶黑暗=14∶10。室外试验在晴天,温度20~30 ℃,湿度50%~60%条件下进行。
1. 2 试验方法
1. 2. 1 雌雄成虫交配前期和雌虫产卵前期观察
选取当天羽化的未交配雄虫和羽化后9~12 d未交配雌虫于种植有九里香的塑料杯内配对,一个塑料杯配一对,每天观察记录雌雄成虫交配情况,直至所有的雄虫均发生交配为止。共配对观察50对成虫。
选取当天羽化的未交配雌虫和羽化后9~12 d未交配雄虫于种植有九里香的塑料杯内配对,一个塑料杯配一对,每天观察记录雌雄成虫的交配情况,直至所有的雌虫均发生交配为止。共配对观察55对成虫。
选取当天羽化的未交配雄虫和雌虫于种植有九里香的塑料杯内配对,一个塑料杯配一对,每天观察记录雌成虫的交配和产卵情况,并统计雌虫自羽化到开始产卵的时间,直至所有的雌虫开始产卵为止。共配对观察50对成虫。
1. 2. 2 交配、产卵和羽化昼夜节律观察 从7:00开始,每1 h观察记录1次室外养虫笼新发生交配的成虫情况,夜间用红光手电筒(4.2 V)协助观察。发现有成虫交配,记录交配时间并统计24 h每个时间段的交配对数,最后统计不同时间段内新发生交配次数占总交配次数的比例。连续观察72 h。
从7:00开始,每1 h观察1次室外养虫笼雌成虫产卵情况,夜间用红光手电筒(4.2 V)协助观察。发现九里香嫩梢处有卵后,将嫩梢剪下,在体视镜下观察并记录不同时段成虫的产卵量,连续观察72 h。最后统计不同时间段内所产下的卵量占总产卵量的比例。
当养虫笼内若虫大量发育到5龄时开始观察24 h中不同时间段若虫羽化的数量,每1 h观察1次,将羽化的成虫取出并在体视镜下观察记录雌雄性别,夜间用红光手电筒(4.2 V)协助观察。最后统计不同时间段内羽化的成虫数量及不同性别成虫的数量。观察到的羽化虫数不应少于200头。
1. 3 统计分析
所有试验数据均采用SPSS 17.0进行统计分析,在GraphPad Prism 5.0下制作图表。
2 结果与分析
2. 1 柑橘木虱雌雄成虫交配和雌虫产卵前期观察结果
观察发现,不同雄虫性发育成熟进度差异较明显,交配前期在7~15 d,平均为9.84±1.79 d(n=50)。从图2可看出,雄虫在羽化后1~6 d均无交配发生,羽化后7 d部分雄虫发育成熟,开始有交尾行为出现,但所占比例仅为8.00%,羽化后第9 d交配比例达最大值,为24.00%。雄虫羽化第13 d时,雄虫累计交配率达96.00%(图3),表明此时多数雄虫已发育至性成熟。
柑橘木虱雌虫的交配前期与雄虫不同,雌虫交配前期在6~16 d,交配前期平均为10.64±2.25 d(n=55)。当天羽化的雌虫不接受交配,羽化后第6 d仅有1.82%的雌虫接受交配,羽化后第9 d雌虫交配比例达最大值,为21.80%,以羽化后9~12 d交配所占比例最大,约占总数的78.14%,部分雌虫在羽化后第16 d才接受交配(图4)。从雌虫累计交配率来看,羽化后第14 d累计交配率达94.41%(图5),表明这一时间段内绝大多数雌虫性成熟,能够与雄虫交配。
柑橘木虱雌虫羽化后需经过9~10 d取食,使卵巢得到发育,第11 d交配后可立即产卵。从图6可知,不同雌性成虫产卵前期的差异明显,最短7 d,最长20 d,平均为12.58±2.90 d(n=50)。雌成虫羽化后一般在第11~12 d进入产卵高峰期,占个体总数的56.00%。
2. 2 柑橘木虱成虫交配、产卵和若虫羽化昼夜节律观察结果
2. 2. 1 成虫交配和产卵昼夜节律 室外观察发现,柑橘木虱成虫的繁殖活动集中发生在3:00—21:00,繁殖活动有一定的昼夜规律,且雌雄成虫活动昼夜节律有明显差异。雌雄成虫交配或产卵前后均需补充营养。21:00—3:00成虫大部分集中在叶背或嫩梢处,除个别成虫发生交配行为外,其余个体一般不活动。
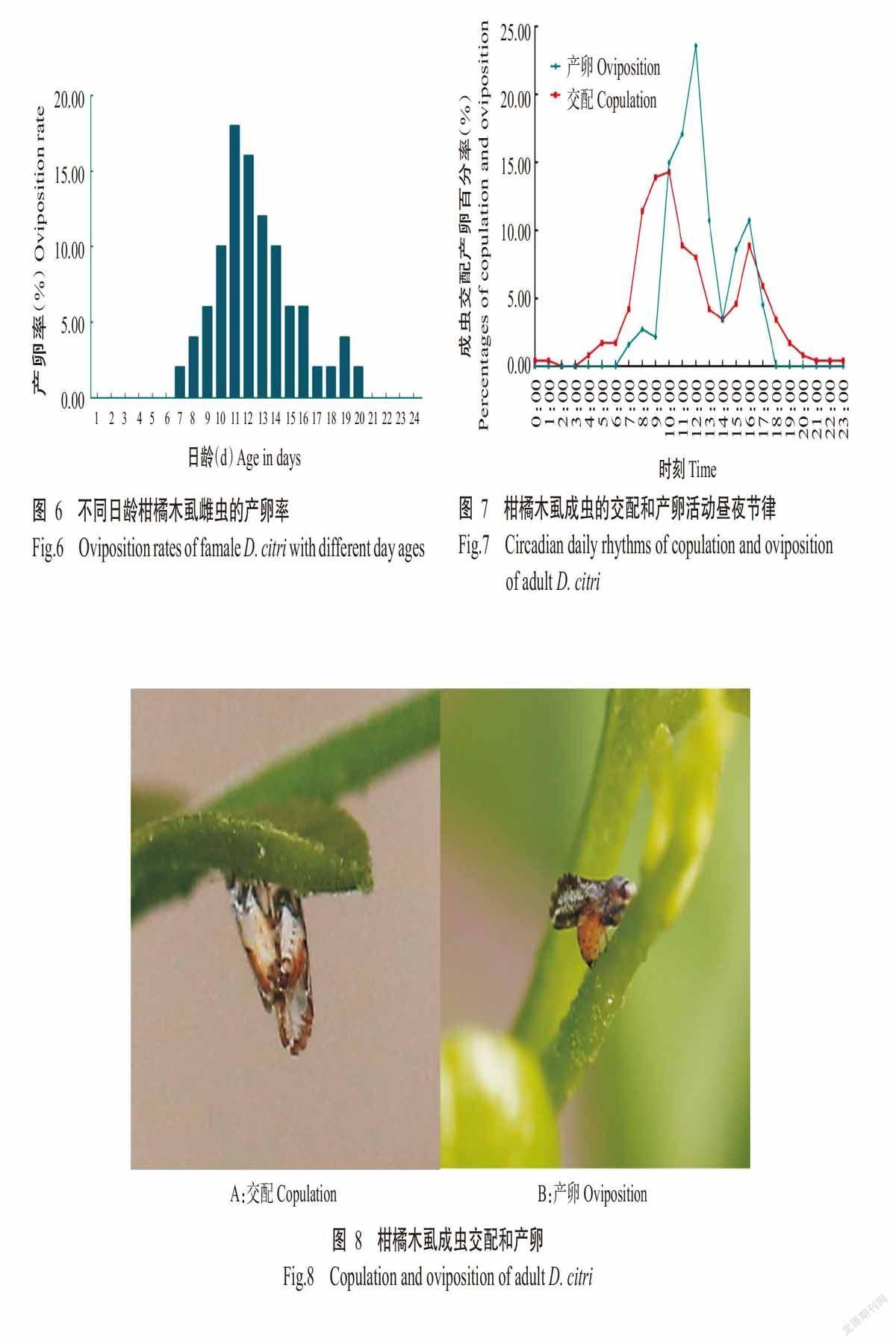
成虫交配在24 h中有两个高峰时段,分别在9:00—10:00和16:00—17:00,其中以9:00—10:00进行交配的个体最多(图7)。在成虫的两个交配高峰时间段内,雄虫求偶和交配活动较活跃,21:00后少有交配发生。雌雄成虫交配时体躯朝同一方向平行排列;雌虫找到合适的产卵地点时,腹部卷曲开始产卵(图8)。
雌虫交配结束后于11:00进入产卵活动期,其产卵有两个高峰时段,分别在11:00—12:00和15:00—16:00,其中以11:00—12:00进行产卵的个体最多(图7)。在雌虫产卵的两个高峰时段,其明显比雄虫活跃,且不接受雄虫的求偶与交配,通常单独在嫩梢顶端寻找合适的部位产卵(图8)。
2. 2. 2 成虫羽化昼夜节律 室外笼养观察发现,羽化前大部分5龄若虫会爬到嫩梢叶或成熟叶片背面停留(图9),开始进入羽化阶段,成虫羽化后在羽化地点直接取食或爬行到植株合适的位置取食。初羽化的成虫大多集中在成熟叶片背面。若虫羽化主要发生在2:00—20:00,20:00—2:00未发现若虫有羽化活动(图10)。
雌雄成虫的羽化高峰时段略有不同。雌虫羽化通常有2个高峰时段,主要集中在9:00—10:00和13:00—14:00,以13:00—14:00羽化的个体数最多;雄虫羽化只有1个高峰时段,主要集中在9:00—10:00(图10)。表明雌雄成虫羽化日节律存在两性差异现象。
3 讨论
本研究结果显示,柑橘木虱雌雄成虫的交配前期分别为10.64±2.25和9.84±1.79 d,雌虫的产卵前期为12.58±2.90 d,说明成虫初羽化都需经历一段时间的取食和性成熟期才能进行交配和产卵,交配和产卵前期是柑橘木虱成虫进行繁殖活动必须经历的阶段,且成虫交配前期存在两性差异,雄成虫的性成熟时间早于雌成虫。谢秀挺等(2016)研究证实柑橘木虱成虫存在繁殖行为前期阶段,且会受温度变化的影响,但该研究并未明确指出成虫的平均交配和产卵前期。部分种类的昆虫如眉斑并脊天牛[Glenea. cantor(Fabricius)]两性羽化出木后均有一个性成熟期才能进行繁殖活动,也证明包括柑橘木虱在内的部分种类昆虫存在一定时长的交配和产卵前期阶段(Wang et al.,1998;Lu et al.,2013)。而扁木虱科昆虫在成虫初羽化后有一段时间的取食发育才能进行繁殖活动,如中国梨木虱[Psylla pyrisuga(Forster)]的产卵前期为6 d(李庆和蔡如希, 1994),其繁殖期早于柑橘木虱成虫,由此可知,不同扁木虱科的昆虫繁殖行为前期存在差异,且扁木虱科昆虫会根据自身性器官发育情况选择在合适的时间进行繁殖活动,也是昆虫进化过程中适应环境的结果。肖培彬等(2017)通过对不同生长阶段柑橘木虱成虫生殖器官的解剖发现,交配前期阶段雄虫的储精囊小且外呈浅黄色,雌虫卵巢和受精囊呈白色,受精囊半透明状,呈不饱满状态;交配期阶段雄虫贮精囊外层变为橙色,明显大于交配前期的贮精囊,雌虫受精囊明显变大;产卵阶段雌虫卵巢内有大量成熟的橙黄色卵,受精囊内有白色精包,证明柑橘木虱成虫在交配前期阶段不具备进行繁殖活动的能力,与本研究结论一致。本研究在室内环境条件下得出柑橘木虱雌雄成虫的平均交配前期和雌成虫平均产卵前期,今后的防治工作可根据成虫存在交配前期这一特点,在成虫还无法进行繁殖活动时期对其进行集中防治,可有效压低种群的虫口数量,控制种群下一代的发生量。本研究虽然获得一定温度条件下成虫交配和产卵前期数据,但在温度变化的情况下是否会对成虫平均繁殖前期产生影响的问题尚未解决,也可能有其他的环境因素对成虫繁殖前期产生影响,因此还需进行更多试验以明确成虫繁殖前期的时长范围。
当柑橘木虱成虫进入繁殖阶段,繁殖活动会呈现一定的规律,成虫交配、产卵和羽化活动主要在2:00—21:00进行,其活动高峰期分别为9:00—10:00、11:00—12:00和9:00—10:00。成虫的繁殖活动发生在白昼,且若虫羽化为成虫的活动集中在成熟叶片背面,雄虫羽化高峰只发生在上午,而雌虫在上午和下午各有一个羽化高峰,由此说明成虫羽化存在两性差异现象。成虫羽化的两性差异现象存在于部分种类昆虫中,如白星花金龟雌虫先于雄虫羽化(王萍莉等,2018),而部分种类的昆虫不存在这一现象,如黄野螟(王忠等,2018)。目前,已有研究对柑橘木虱成虫繁殖活动的生物学特性进行详细记录(王秀娟,2010),但繁殖活动的昼夜节律问题尚无详细报道。Mankin等(2013)研究指出,昆虫在合适的时间和地点活动可减少被天敌发现的概率,并可减少分享同一资源的不同物种间的相互竞争,在获得充足食料的前提下使昆虫种群的繁殖活动有规律地进行。柑橘木虱成虫繁殖活动昼夜节律现象也是其与生态系统中其他物种竞争生存空间的结果。本研究详细记录了柑橘木虱成虫繁殖活动的昼夜节律,可为今后根据柑橘木虱成虫繁殖行为日节律的特点对其进行防治及预测预报提供参考,但在不同季节成虫的繁殖活动是否也符合这一规律有待进一步探究。
昆虫繁殖特性及活动规律是其长期进化的结果,成虫在环境条件较稳定的情况下其繁殖活动规律不会有明显变化。因此,探明昆虫繁殖前期及繁殖活动规律,可为害虫种群的预测预报提供参考。本研究通过探讨柑橘木虱成虫繁殖特性和繁殖活动昼夜节律,可为今后防治柑橘木虱提供基础数据,最终为从行为学角度防治该害虫提供新方法。
4 结论
柑橘木虱雌雄成虫交配和雌虫产卵前期分别为10.64±2.25、9.84±1.79和12.58±2.90 d,雌雄成虫交配前期存在两性差异;交配、产卵和羽化活动昼夜节律集中在2:00—21:00,各活动高峰期分别为9:00—10:00、11:00—12:00和9:00—10:00,雌雄成虫羽化节律存在两性差异。可根据柑橘木虱成虫繁殖行为前期及活动规律对其进行防治及预测预报。
参考文献:
贾志成,郑加强,黄雅杰,周宏平,Ehsani Reza. 2015. 柑橘黄龙病热处理防治技术研究进展[J]. 农业工程学报,31(23):1-9. [Jia Z C,Zheng J Q,Huang Y J,Zhou H P,Ehsani R. 2015. Review and prospect of thermotherapy for citrus Huanglongbing[J]. Transactions of the Chinese Society of Agricultural Engineering,31(23): 1-9.]
李超峰,劉家莉,曾鑫年. 2019. 柑橘木虱趋光行为及复眼结构分析[J]. 华南农业大学学报, 40(2): 53-59. [Li C F,Liu J L,Zeng X N. 2019. Phototactic behavior and compound eye structure of Diaphorina citri[J]. Journal of South China Agricultural University, 40(2): 53-59.]
李慶,蔡如希. 1994. 温度对梨木虱生长发育的影响[J]. 西南农业大学学报,16(2):175-177. [Li Q,Cai R X. 1994. Effects of temperature on the development of pear leaf psylla(Psylla pyrisuga)[J]. Journal of Southwest Agricultural University, 16(2): 175-177.]
王萍莉,李小万,高朋,温俊宝. 2018. 白星花金龟的羽化及交配行为[J]. 植物保护,44(1):174-178. [Wang P L,Li X W,Gao P,Wen J B. 2018. The emergence and mating behavior of Protaetia brevitarsis Lewis[J]. Plant Protection, 44(1):174-178.]
王秀娟. 2010. 柑橘木虱与两种寄主植物互作的研究[D]. 福州:福建农林大学. [Wang X J. 2010. Interactions between Diaphorina citri Kuwayama and its two species of host plants[D]. Fuzhou: Fujian Agricultural and Fores-try University.]
王忠,谢伟忠,朱诚棋,陆雪雷,曹春雷,温修军. 2018. 黄野螟的羽化和生殖行为节律[J]. 中国森林病虫,37(1):24-30. [Wang Z,Xie W Z,Zhu C Q,Lu X L,Cao C L,Wen X J. 2018. Circadian rhythm of emergence and reproduction of Heortia vitessoides Moore(Lepidoptera:Crambidae)[J]. Forest Pest and Disease, 37(1):24-30.]
肖培彬,马义雷,李皓,韩群鑫. 2017. 亚洲柑橘木虱内生殖系统形态变化规律研究[J]. 环境昆虫学报, 39(6):1207-1213. [Xiao P B,Ma Y L,Li H,Han Q X. 2017. Morphological changes of the internal reproductive system of Diaphorina citri Kuwayama[J]. Journal of Environmental Entomology, 39(6): 1207-1213.]
谢秀挺,刘卫东,彭龙,张倩,方贻文,严翔,李梦媛,张丽艳,赖华荣. 2016. 柑桔木虱成虫产卵习性研究[J]. 中国南方果树,45(2): 69-71. [Xie X T,Liu W D,Peng L,Zhang Q,Fang Y W,Yan X,Li M Y,Zhang L Y,Lai H R. 2016. Study on the oviposition of citrus psyllid adult[J]. South China Fruits,45(2):69-71.]
Brar G S,Meyer W,Stelinski L L. 2015. Effects of methoprene,a juvenile hormone analog,on survival of various developmental stages,adult emergence,reproduction, and behavior of Asian citrus psyllid,Diaphorina citri Kuwayama[J]. Pest Management Science,71(12):1657-1665.
Erikj W,Davidg H,Richardw M. 2009. Vibrational Communication Between the Sexes in Diaphorina citri(Hemiptera: Psyllidae)[J]. Annals of the Entomological Society of America, 102(3): 547-555.
Ibanez F,Racine K,Hoyte A,Stelinski L L. 2019. Reproductive performance among color morphs of Diaphorina citri Kuwayama, vector of citrus greening pathogens[J]. Journal of Insect Physiology,117:103904.
Kelley A J,Pelz-Stelinski K S. 2019. Maternal contribution of Candidatus Liberibacter asiaticus to Asian citrus psyllid (Hemiptera:Liviidae) nymphs through oviposition site inoculation and transovarial transmission[J]. Journal of Economic Entomology,10:1-4.
Lu W,Wang Q,Tian M Y,Xu J,Lv J,Wei S G,Qin A Z. 2013. Reproductive traits of Glenea cantor(Coleoptera: Cerambycidae:Lamiinae)[J]. Journal of Economic Entomology,106(1):215-220.
Mankin R W,Rohde B B,Mcneill S A,Paris T M,Zagvazdina N I,Greenfeder S. 2013. Diaphorina citri(Hemiptera: Liviidae) responses to microcontroller-buzzer communication signals of potential use in vibration traps[J]. Florida Entomologist,96(4): 1546-1555.
Onagbola E O,Meyer W L,Boina D R,Stelinski L L. 2008. Morphological characterization of the antennal sensilla of the Asian citrus psyllid, Diaphorina citri Kuwayama(Hemiptera: Psyllidae),with reference to their probable functions[J]. Micron,39(8):1184-1191.
Qasim M,Lin Y,Dash C K,Bamisile B S,Ravindran K,Islam S U,Ali H,Wang F,Wang L. 2019. Temperature-dependent development of Asian citrus psyllid on various hosts,and mortality by two strains of Isaria[J]. Microbial Pathogenesis,119:109-118.
Rwomushana I,Khamis F M,Grout T G,Mohamed S A,Sétamou M,Borgemeister C,Heya H M,Tanga C M,Nderitu P W,Seguni Z S,Materu C L,Ekesi S. 2017. Detection of Diaphorina citri Kuwayama(Hemiptera:Liviidae) in kenya and potential implication for the spread of huang-longbing disease in East Africa[J]. Biological Invasions,19(10):2777-2787.
Stockton D G,Martini X,Stelinski L L. 2017. Male psyllids differentially learn in the context of copulation[J]. Insects,8: 1-14.
Wang Q,Shi G L,Davis L K. 1998. Reproductive potential and daily reproductive rhythms of Oemona hirta(Coleoptera:Cerambycidae)[J]. Journal of Economic Entomology,91(6): 1360-1365.
(責任编辑 麻小燕)
猜你喜欢
电脑报(2022年24期)2022-07-01
新农民(2021年21期)2021-09-17
华夏地理(2019年2期)2019-07-24
植物保护(2018年1期)2018-05-14
小溪流(画刊)(2017年6期)2017-07-10
电脑知识与技术(2016年32期)2017-03-17
理科考试研究·高中(2016年10期)2017-01-17
理科考试研究·高中(2016年5期)2016-05-14
江苏农业科学(2015年1期)2015-04-17
少年科学(2009年12期)2009-07-07
