
东方田鼠感染日本血吸虫前后血常规和血清生化指标的动态变化
2019-09-06 01:27:28柴淑梅余新刚林矫矫柏熊谢建芸傅志强
中国动物传染病学报 2019年4期
柴淑梅 ,张 倩 ,翟 颀 ,余新刚 ,洪 炀 ,陆 珂 ,李 浩 ,林矫矫 ,柏熊 ,谢建芸,傅志强
(1.中国农业科学院上海兽医研究所 农业部动物寄生虫学重点开放实验室,上海 200241;2.上海实验动物研究中心,上海 201203)
日本血吸虫病(schistosomiasis japonicum)是一种危害严重的人畜共患寄生虫病。日本血吸虫(Schistosoma japonicum,S. japonicum)是该病病原,可寄生于7属41种哺乳动物,不同种类的宿主对日本血吸虫感染的适宜性差异较大[1]。根据血吸虫在感染宿主体内的生长发育情况,可将宿主分为适宜宿主和非适宜宿主。东方田鼠(Microtus fortis,M.fortis)是我国迄今为止发现的唯一一种具有天然抗日本血吸虫病特征的哺乳动物。日本血吸虫在东方田鼠体内先后经历皮肤期童虫、肺期童虫及肝期童虫的发育过程,最终在肝脏消亡[2]。多位学者从血清、组织蛋白质组学和差异基因等多个途径研究了东方田鼠与其他宿主之间差异,然而东方田鼠天然抗日本血吸虫的机制仍然是一个未解之谜[3-4]。
血吸虫尾蚴侵入终末宿主皮肤后通过毛细血管和毛细淋巴管进入循环系统,此后在其中生长发育。血液不仅为虫体生长发育提供营养物质,也是宿主对其产生免疫应答的主要环境,因此分析血液中的免疫细胞和生化物质是了解宿主和血吸虫相互作用的重要途径。邵伟娟等[5-6]测定了东方田鼠正常生理状态下的血液和生化血清指标,结果显示部分指标与小鼠有差异。本研究比较分析了BALB/c鼠、东方田鼠未感染和感染日本血吸虫后不同时间点的血常规和生化各项指标变化情况。
1 材料与方法
1.1 生物材料清洁级东方田鼠长江亚种(70 g,雄性)由上海实验动物研究中心提供;BALB/c鼠(25 g,雄性)购于上海斯莱克实验动物有限责任公司;日本血吸虫中国大陆株尾蚴由中国农业科学院上海兽医研究所动物血吸虫病研究室提供。
1.2 主要仪器血细胞分析仪(迈瑞BC5300),生化分析仪(迈瑞BS200),TOMY全自动高压灭菌锅(SX-500,日本TOMY公司),纯水/超纯水一体化系统(Millipore公司)。
1.3 血常规检测东方田鼠、BALB/c鼠各20只,分别腹部攻击感染日本血吸虫尾蚴100条,并在感染后第0、1、3、5、8、12、16、21、28、35、42 d进行尾部毛细管采血 20 μL。将采集血液迅速转移到含180 μL稀释液的1.5 mL离心管中吹打混匀,放置2 min 后采用血细胞分析仪检测。检测指标包括:白细胞(white blood cell,WBC)、嗜中性粒细胞(neutrophils,Neu)、淋巴细胞(lymphocyte,Lym)、单核细胞(monocytes,Mon)、嗜酸性粒细胞(eosinophil,Eos)、嗜碱性粒细胞(basophil,Bas)、红细胞(red blood cell,RBC)、血红蛋白(hemoglobin,HGB)、血细胞压积(packcd cell volume,HCT)、红细胞平均容量(mean corpuscular volume,MCV)、红细胞平均血红蛋白容量(mean corpuscular hemoglobin,MCH)、红细胞平均血细胞浓度(mean corpuscular hemoglobin concentration,MCHC)和血小板计数(platelet,PLT)。所有样本均在2 h内检测完毕。
1.4 血清生化检测东方田鼠、BALB/c鼠各40只,分别腹部攻击感染日本血吸虫尾蚴100条,并在感染后第0、1、3、5、8、12、16、21、28、35、42 d进行摘眼球采血并处死。全血室温下静置30 min,4℃静置30 min,400×g离心15 min,收集血清-20℃保存备用。生化指标包括总蛋白(total protein,TP)、白蛋白(albumin,ALB)、C-反应蛋白(C-reactive protein,CRP)、前白蛋白(prealbumin,PA)、高密度脂蛋白胆固醇(highdensity lipoprotein cholesterol,HDL-C)、低密度脂蛋白胆固醇(low density lipoprotein cholesterol,LDL-C)、载脂蛋白A1(apolipoproteinⅠ,ApoA1)、免疫球蛋白G(immunoglobulin G,IgG)、免疫球蛋白M(immunoglobulin M,IgM)、补体C3 (complement C3)和补体C4(complement C4)等。
2 结果
2.1 血细胞变化血常规检测东方田鼠感染血吸虫前后血细胞含量及其变化(表1)发现,东方田鼠感染日本血吸虫尾蚴后第1~16 d期间WBC、Neu、Lym、Mon、Eos、Bas等显著高于感染前(0 d),BALB/c鼠感染后血象变化稍有差异,但不显著(表2)。比较小鼠与东方田鼠的白细胞分类含量及其变化发现,东方田鼠感染日本血吸虫尾蚴第1~16 d 期间WBC显著高于正常值,主要原因是Neu的显著升高;而小鼠感染血吸虫后WBC数量升高,但只有第5、16 d显著高于感染前(0 d),且在正常参考值之内,可能与Eos和Lym的升高相关(图1)。

图1 东方田鼠、BALB/c鼠感染日本血吸虫后白细胞及分类细胞变化趋势Fig.1 The dynamic changes of white blood cell and its classi fication from S. japonicum-infected Microtus fortis and BALB/c mouse
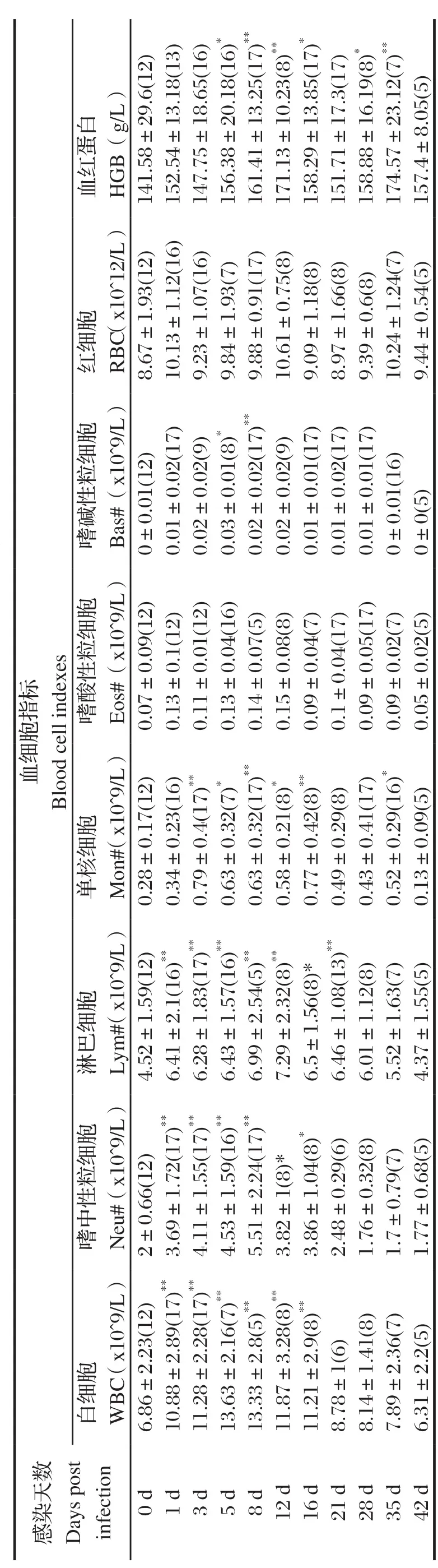
表1 东方田鼠感染日本血吸虫前后血细胞指标Table 1 Blood cell indexes of Microtus fortis before and after infection of Schistosoma japonicum
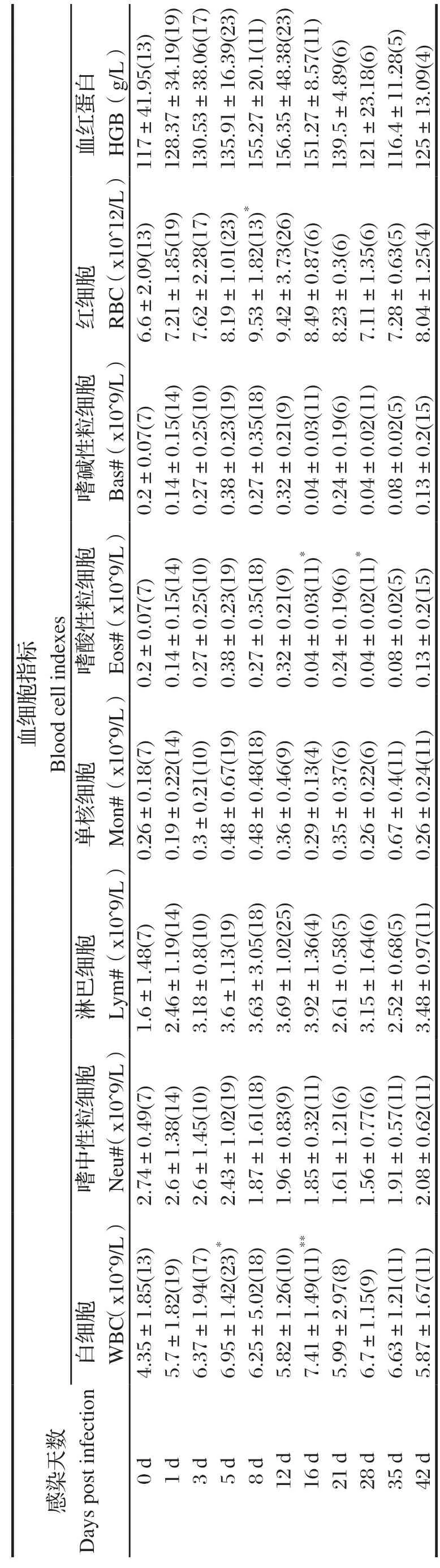
表2 BALB/c 小鼠感染日本血吸虫前后血细胞指标Table 2 Blood cell indexes of BALB/c mice befor e and after infection of Schistosoma japonicum
2.2 血清生化变化东方田鼠、BALB/c鼠感染日本血吸虫尾蚴前后血清生化指标含量及其结果见表3~4。东方田鼠感染后ALB、HDL-C显著升高,BALB/c鼠IgG、IgM在感染后第4周显著升高,其他指标变化不显著。比较东方田鼠和小鼠的各指标结果表明,东方田鼠血清TP、LDL-C、ApoA1、PA含量高于小鼠,IgM、HDL-C含量低于小鼠(图2)。

表3 东方田鼠感染日本血吸虫前后血清生化指标Table 3 Blood biochemical indexes of Microtus fortis before and after infection of Schistosoma j
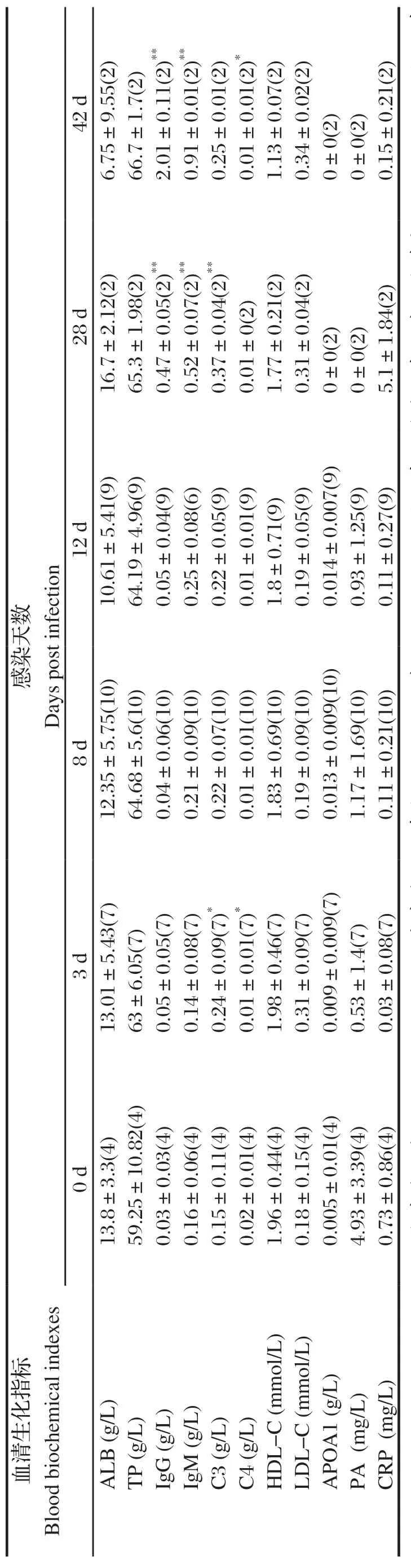
表4 BALB/c 小鼠感染日本血吸虫前后血清生化指标Table 4 Blood biochemical indexes of BALB/c mice befor e and after infection of Schistosoma japonicum
3 讨论
日本血吸虫可自然寄生于40多种哺乳动物,唯独不能在东方田鼠中发育成熟,这一独特的生物学现象及抗性机制引起了广泛关注。宿主和寄生虫的相互作用是寄生虫能否完成生活史和宿主发病情况的重要因素[7]。东方田鼠对日本血吸虫的抗性为研究宿主杀伤血吸虫,控制和限制感染的宿主-寄生虫相互作用提供了良好的模型系统。血吸虫尾蚴侵入东方田鼠皮肤后进入其循环系统,血液是血吸虫童虫赖以生存摄取营养的主要环境,其中的细胞和生化物质可能是其抵抗感染的重要因素,因此分析东方田鼠感染日本血吸虫尾蚴后血细胞和生化物质的含量及其变化可为抗性机制的研究提供线索。
天然免疫细胞是参与先天性免疫应答的免疫细胞统称,包括吞噬细胞(单核巨噬细胞、中性粒细胞、嗜酸性粒细胞、嗜碱性粒细胞等)和非吞噬性先天性免疫细胞(NK、NKT等)。这些细胞在先天性免疫应答中起重要作用,也可通过分泌细胞因子调节适应性免疫应答[8-9]。蒋伟斌等[10]表明,东方田鼠感染日本血吸虫后第3~16 d血液嗜酸性粒细胞具有趋化性和粗大的嗜酸性颗粒,颗粒内含有过氧化物酶和酸性磷酸酶,能有效的杀灭寄生虫,刘金明等[11]发现东方田鼠EOS能粘附在日本血吸虫童虫表面,是否起到杀虫作用仍然未知。胡媛等[12]在东方田鼠免疫抑制模型中成功抑制了淋巴细胞的增值,但东方田鼠对日本血吸虫感染的表型没有改变,推测淋巴细胞在东方田鼠抗血吸虫病机制中可能不发挥直接作用。本研究分析比较了东方田鼠和BALB/c鼠感染日本血吸虫后血常规五分类的含量和动态变化,结果表明东方田鼠外周血中Neu的含量显著大于BALB/c鼠的正常参考值,而且其变化趋势与感染过程有一定的相关性。中性粒细胞具有变形运动和吞噬活动的能力,细胞表面具有补体和免疫球蛋白受体,是机体对抗病原最重要的防卫系统[13]。提示Neu可能在东方田鼠抗日本血吸虫机制发挥了重要作用,可能与补体杀伤日本血吸虫机制相关。

图2 东方田鼠、BALB/c鼠感染日本血吸虫后血清生化指标变化趋势Fig.2 The dynamic changes of serum biochemical indexes from S. japonicum-infected Microtus fortis and BALB/c mouse
总蛋白和白蛋白含量及变化是机体蛋白质代谢的敏感标识[14],也可反映机体的免疫功能状况。血清白蛋白与球蛋白的比值可以反映机体免疫系统状态,如果在适当范围内球蛋白合成提高或白蛋白合成下降,机体免疫功能就会有增强趋势。有研究表明,血吸虫可以利用宿主的白蛋白作为抗氧化剂,是虫体在宿主体内生存必需的物质[15-16],Holtfreter等[17]研究表明,宿主的白蛋白是血吸虫童虫在早期发育过程中的主要能量来源,也是成虫的辅助能量来源。邵伟娟等[18]研究发现东方田鼠正常生理状态下血清白蛋白与球蛋白比例倒置。Li等[19]研究表明东方田鼠血清ALB与日本血吸虫童虫孵育可以诱导46.2%的童虫死亡率,显著高于阴性对照;与对照组动物相比,注射Mf-白蛋白的小鼠可诱导43.5%的减虫率和48.1%的减卵率。本研究结果显示东方田鼠感染日本血吸虫尾蚴后血清ALB含量显著增加。要进一步明确白蛋白在东方田鼠抗日本血吸虫特性中的作用还需要更深入的研究。血清高密度脂蛋白质(high density lipoprotein,HDL)是肝病预后判断的重要指标,与肝病病情严重程度密切相关,HDL降低是病情恶化的表现,当患肝硬化或重型肝炎时,其含量会下降[20]。但是,HDL 可随着病情的好转而逐渐升高。HDL-C是临床检验的指标,它代表了血液中HDL的水平。东方田鼠感染日本血吸虫后HDL-C不降反而逐渐升高,说明其逐渐自愈血吸虫带来的组织损伤。
猜你喜欢
作文大王·低年级(2021年11期)2021-12-22 00:58:13
小天使·一年级语数英综合(2021年2期)2021-05-08 07:48:34
热带病与寄生虫学(2021年2期)2021-02-28 15:15:11
科学(2020年3期)2020-11-26 08:18:22
当代水产(2020年3期)2020-06-15 12:03:02
小学生(看图说画)(2020年4期)2020-04-01 11:58:18
中国感染与化疗杂志(2020年1期)2020-01-12 14:07:30
实用皮肤病学杂志(2015年4期)2015-12-22 11:21:42
吉林大学学报(医学版)(2015年1期)2015-12-17 07:47:23
医学研究杂志(2015年12期)2015-06-10 06:57:46
