
甘蓝型油菜SNARE蛋白SYP122叶片瞬时表达体系的建立
2019-09-05 09:20:16单亚楠陈淞渝王爱荣
福建农林大学学报(自然科学版) 2019年4期
许 静, 单亚楠, 何 豆, 陈淞渝, 庄 炜, 王爱荣
(福建农林大学植物保护学院,福建 福州 350002)
油菜是我国继水稻、玉米、小麦、大豆之后的第五大作物[1],种植面积超过750万hm2[2],其不仅是重要的油料作物,还是潜在的生物能源原料[3].油菜主要有3个栽培种,即甘蓝型油菜(Brassicanapus)、芥菜型油菜(B.juncea)和白菜型油菜(B.rapa)[4],其中以甘蓝型油菜的种植范围最广[5].
随着基因工程技术的迅猛发展,转基因技术成为油菜基因功能研究的重要方法.目前常用的遗传转化方法有根癌农杆菌介导法、聚乙二醇(polyethylene glycol, PEG)法、基因枪法、真空渗透遗传转化法和花粉介导法等[6-7].这些方法都不可避免地要经过脱分化及再分化过程,不仅耗时长、影响因素众多,而且对操作者的技术能力及试验条件都有较高的要求.农杆菌介导的基因瞬时表达系统可在短时间内大量表达外源基因,不需要经过世代培养,这使其成为基因功能研究的利器[8].近年来,学者们虽然针对油菜建立了农杆菌介导的瞬时表达系统,但是仅局限于子叶[6]和原生质体[9].为了高效地开展油菜基因功能的研究,在油菜叶片中建立一个成熟的基因表达体系显得尤为重要.
植物SNARE(soluble N-ethylmaleimide-sensitive factor attachment protein receptors)蛋白是参与细胞膜融合的一类重要蛋白,其家族成员在多种生物学过程中发挥重要作用,包括生长动态平衡、激素调节、胁迫反应和免疫反应[10-12].这类蛋白中,研究最深入的是PEN1(SYP121),它对植物抗白粉病起着关键作用[10].拟南芥(Arabidopsisthaliana)AtSYP121及其同源蛋白AtSYP122在植物的防御信号途径中起着负调控作用,是细胞程序性死亡(programmed cell death, PCD)以及水杨酸(salicylic acid, SA)、茉莉酸(jasmonic acid, JA)和乙烯(ethylene, ET)信号通路的负调控因子[13].然而,AtSYP122对植物抗白粉病的作用很小[14-15].在细胞膜分泌或锚定在细胞膜的蛋白中,二者的货物蛋白群体也存在较大差别[16-17].本实验室前期研究发现,油菜SYP122同源蛋白在抗死体营养型病原菌——核盘菌(Sclerotiniasclerotiorum)方面发挥重要功能.
本研究拟克隆甘蓝型油菜中双九号的BnSYP122基因,并将其构建到带有黄色荧光蛋白(yellow fluorescent protein, YFP)的植物表达载体pEarleyGate 104上,通过农杆菌介导的真空渗透法在4周龄油菜的叶片中表达YFP-BnSYP122及绿色荧光蛋白(green fluorescent protein, GFP),通过荧光观察和Western blot 检测分析其表达效果,初步验证油菜BnSYP122的亚细胞定位,以期为进一步阐明BnSYP122在油菜抗菌核病中的功能提供一定的依据,并为油菜基因功能研究提供一定的技术支持.
1 材料与方法
1.1 材料
供试植物为甘蓝型油菜中双九号,种子播于含黑土和蛭石(1∶1)的营养土中,置于23 ℃的温室中,光暗比为16 h∶8 h,光照度为5 500 lx,覆盖保鲜膜培养3~4 d.待幼苗子叶展开,去掉保鲜膜,一周后将油菜苗移栽至新的花盆中,每3~5 d浇水一次,10 d浇营养液一次,继续培养3~4周,至油菜长出4~5片真叶,即可用于瞬时表达.
瞬时表达所用的对照载体pEGAD-GFP、构建重组载体所用Gateway方法克隆入门载体pDONR/Zeo、植物表达载体pEarleyGate 104(35S-YFP-Gateway-OCS3′)为本实验室留存.试验所用的真空泵为AF2000真空抽滤器(购自宁波奉化晨光威腾自动化机械有限公司).
1.2 方法
1.2.1BnSYP122基因的鉴定及序列分析 在拟南芥数据库(https://www.arabidopsis.org/)中搜索AtSYP122,得到其编码基因及蛋白的氨基酸序列,将该序列作为模板在油菜数据库(http://www.genoscope.cns.fr/brassicanapus/)中BLAST之后,获得与AtSYP122最同源的序列,根据该序列的开放阅读框(open reading frame, ORF)设计引物.采用油菜中抗品种中双九号[18]的cDNA作为模板进行PCR扩增,将获得的片段构建于pMD18-T载体上,送上海生工生物科技有限公司测序.将获得的序列翻译后与拟南芥AtSYP122进行比对分析.
将扩增获得的AtSYP122在油菜中的同源基因序列翻译成氨基酸序列后,在SMART(http://smart.embl-heidelberg.de/)中分析其保守功能域;用在线软件SOPMA(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=%2FNPSA%2Fnpsa_seccons.html)预测分析其二级结构;同时,在http://www.cbs.dtu.dk/services/TMHMM-2.0/中进行跨膜结构域的预测.
1.2.2 表达载体的构建 根据Gateway试剂盒说明书设计引物,两端加上接头attB,以油菜cDNA为模板扩增获得油菜BnSYP122基因全长.按照Gateway试剂盒说明书将扩增到的PCR产物通过BP反应连接到pDONR/Zeo载体,并转化DH5α感受态,涂布于含Zeocin的LB培养基,37 ℃倒置培养,20 h后挑选单克隆进行PCR验证.将验证正确的转化子送上海生工生物科技有限公司测序.选择序列正确的单克隆,提取质粒进行LR反应,同样转化DH5α感受态并筛选转化子,将验证正确的转化子进一步采用冻融法转化农杆菌GV3101菌株.
1.2.3 真空渗透法表达目的蛋白 挑取农杆菌单克隆置于含有5 mL LB液体培养基和终浓度为50 ng·mL-1卡那霉素的试管中,28 ℃、200 r·min-1振荡培养36 h后,以1∶1 000的比例转接至含有150 mL LB液体培养基和终浓度为50 ng·mL-1卡那霉素的锥形瓶中,继续培养至D600 nm为0.6~0.8,离心并收集菌体.然后用渗透缓冲液(10 mmol·L-1MgCl2,10 mmol·L-1MES,150 μmol·L-1乙酰丁香酮,0.02% Silwet L-77)重悬,离心并重悬3次后,调节D600 nm为0.6,28 ℃静置2~3 h.
将农杆菌悬浮液置于广口瓶或培养盒中与油菜并排放置在AF2000真空抽滤器中,油菜的叶片向下浸入到农杆菌悬浮液中,打开真空泵,设置压力最大值为0.06 MPa,保持真空状态15~20 min.取出油菜植株并用吸水纸将叶片表面水分沾干,可观察到50%油菜叶片出现水渍斑,黑暗条件下放置12 h后,移到光照条件下正常培养2~3 d.
1.2.4 激光共聚焦显微镜观察目的蛋白的表达情况 用刀片切取3 mm×3 mm大小的侵染过农杆菌的油菜叶片,下表皮朝上置于载玻片,盖上盖玻片并在盖玻片与载玻片之间滴一滴无菌水,置于激光共聚焦显微镜下,在488 nm激光下观察.
1.2.5 Western blot验证目的蛋白的表达情况 取200 mg油菜叶片于液氮中充分研磨,转移到2 mL EP管中并加入100 μL蛋白提取缓冲液[50 mmol·L-1Tris (pH 7.5),150 mmol·L-1NaCl,0.5% Triton X-100,0.5% Nonidet P-40,0.25% Na-deoxycholate, 1×蛋白酶抑制剂],充分涡旋混匀.12 000 r·min-1离心15 min,吸取上清液,等体积加入2×SDS蛋白上样缓冲液,煮沸10 min变性.
取20 μL蛋白样品跑SDS-PAGE胶,进行Western blot检测,转膜后按照1∶5 000的比例加入GFP羊抗鼠一抗进行杂交,随后以1∶8 000的比例加入辣根过氧化物酶(horseradish peroxidase, HRP)标记的二抗,将杂交后的膜加入western BrightTMECL显影剂后在化学发光成像仪tanon-5200下观察.
2 结果与分析
2.1 油菜BnSYP122蛋白的鉴定
以拟南芥AtSYP122的氨基酸序列为种子序列在已公布的油菜蛋白数据库中进行BLAST搜索,结果比对到与其同源性最高的蛋白序列BnaA04g05470D.根据此蛋白的基因序列设计引物,以油菜中双九号的cDNA为模板进行片段扩增并测序,经翻译后将其蛋白序列BnSYP122_ZS9与数据库中的BnaA04g05470D、AtSYP122进行多重序列比对分析.结果发现,扩增基因的蛋白序列与BnaA04g05470D存在4个氨基酸的差别(图1).但是,这2个蛋白序列与AtSYP122的同源性高达80.4%,并且都具有保守的SNARE 结构域(SNARE-domain)(图1).这表明鉴定和扩增获得的蛋白是典型的SNARE蛋白,可能与AtSYP122具有类似的功能,并将扩增获得的蛋白序列命名为BnSYP122.

黑色填充部分表示氨基酸序列完全相同;蓝色填充部分表示氨基酸序列有2个相同;黑色横线区域为t-SNARE保守基序;红色箭头指示BnSYP122_ZS9与BnA04g05470D的差异氨基酸.图1 BnSYP122和AtSYP122的序列比对Fig.1 Sequence alignment of BnSYP122 and AtSYP122
2.2 BnSYP122蛋白结构预测与分析
SMART预测表明,BnSYP122 N端的39~165位氨基酸为SynN结构域(图2A、2B);通过SOPMA对其二级结构进行预测发现,在蛋白的N端依次排列着Ha﹑Hb和Hc 3个α-螺旋束(图2D).这表明BnSYP122是SNARE蛋白家族的突触融合(syntaxin)蛋白.在其C端的286~305位存在一个跨膜结构域(图2A、2B),表明BnSYP122可能为跨膜蛋白,定位于细胞膜.为了确认其可靠性,使用TMHMM 2.0再次预测了BnSYP122的跨膜结构域(图2C),该结果与SMART的预测结果一致.

A.SMART预测的结构域示意图;B.SMART预测的结构域列表;C.TMHMM 2.0预测的跨膜结构示意图;D.SOPMA预测的N端二级结构示意图.图2 BnSYP122的结构域分析Fig.2 Structure analysis of BnSYP122
2.3 表达载体的构建与农杆菌转化
以油菜中双九号的cDNA为模板进行PCR扩增,产物经1%琼脂糖凝胶电泳,检测到约1 000 bp的目的条带(图3A),切胶回收后进行Gateway入门反应,并转化大肠杆菌DH5α感受态.将Zeocin筛选的单克隆,以pDONR载体引物M13-F搭配片段引物BnSYP122-R进行PCR验证,得到略大于1 000 bp的条带(图3B).测序正确后提取质粒与载体pEarleyGate 104进行LR反应,转化大肠杆菌DH5α感受态,挑选经卡那霉素筛选后的单克隆进行菌落PCR验证,4个单克隆均扩增到约1 000 bp的条带(图3C).提取质粒采用冻融法转化农杆菌GV3101,卡那霉素和利福平筛选到的单克隆经PCR验证为阳性克隆(图3D).以上结果表明, pEarleyGate 104-BnSYP122重组载体构建成功,并转化获得农杆菌菌株,可用于下一步的油菜侵染试验.
2.4 油菜叶片目的蛋白的荧光分析
为了验证GFP和目的蛋白BnSYP122是否在油菜叶片中正常表达,使用激光共聚焦显微镜分别观察用真空渗透法表达GFP和YFP-BnSYP122的油菜叶片.如图4B所示,在处理的叶片细胞中可以观察到均匀分布的绿色荧光,表明外源基因可以在油菜叶片中正常翻译成蛋白.将BnSYP122蛋白融合YFP标签进行定位观察,结果显示,与位于细胞膜、细胞质及细胞核的GFP相比,BnSYP122明显定位于细胞膜(图4C),与生物信息学预测结果一致.以上结果说明真空渗透法不仅能够在油菜叶片中成功表达目的蛋白,还能够准确显示目的蛋白的亚细胞定位.
2.5 Western blot验证目的蛋白的表达
为了进一步确定目的蛋白在油菜叶片中的正常表达,收集经含有GFP和YFP-BnSYP122表达载体的根癌农杆菌菌液真空渗透处理2 d的油菜叶片,提取总蛋白,用Western blot检测其蛋白表达量.结果显示,用GFP抗体杂交后获得与目的蛋白大小一致的条带(图5),说明真空渗透法确实能够正常表达目的蛋白.
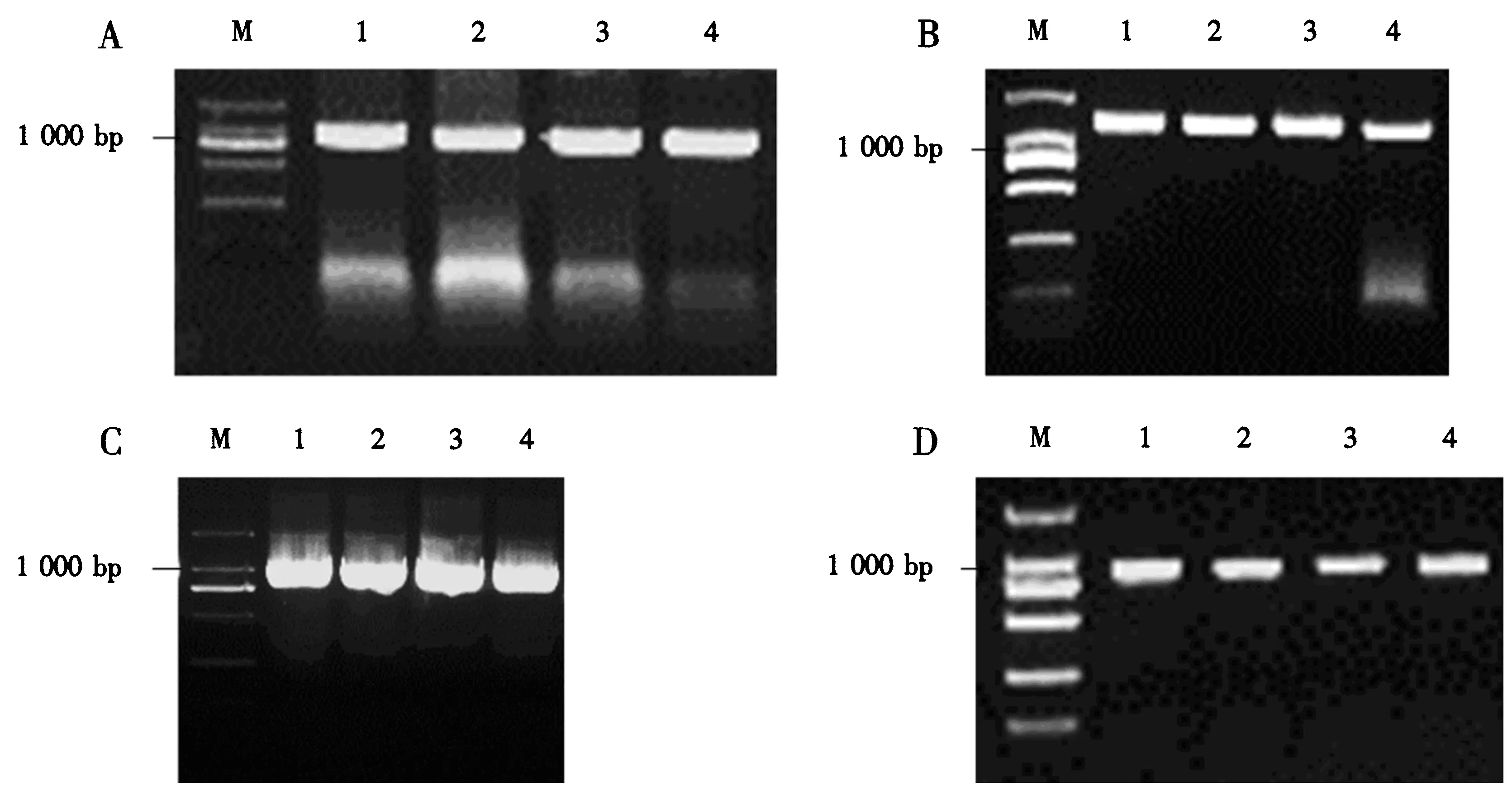
A.BnSYP122的扩增,其中M为Marker DL2000,1-4为扩增到的PCR片段;B.pDONR/Zeo-BnSYP122菌落PCR验证,其中M为Marker DL2000,1-4为大肠杆菌单克隆;C.pEarleyGate 104-BnSYP122菌落PCR验证,其中M为Marker DL2000,1-4为大肠杆菌单克隆;D.菌落PCR验证GV3101转化子,其中M为Marker DL2000,1-4为农杆菌单克隆.图3 BnSYP122的扩增及Gateway法构建重组载体的验证Fig.3 Amplification of BnSYP122 and verification of recombinant vector constructed by Gateway technique
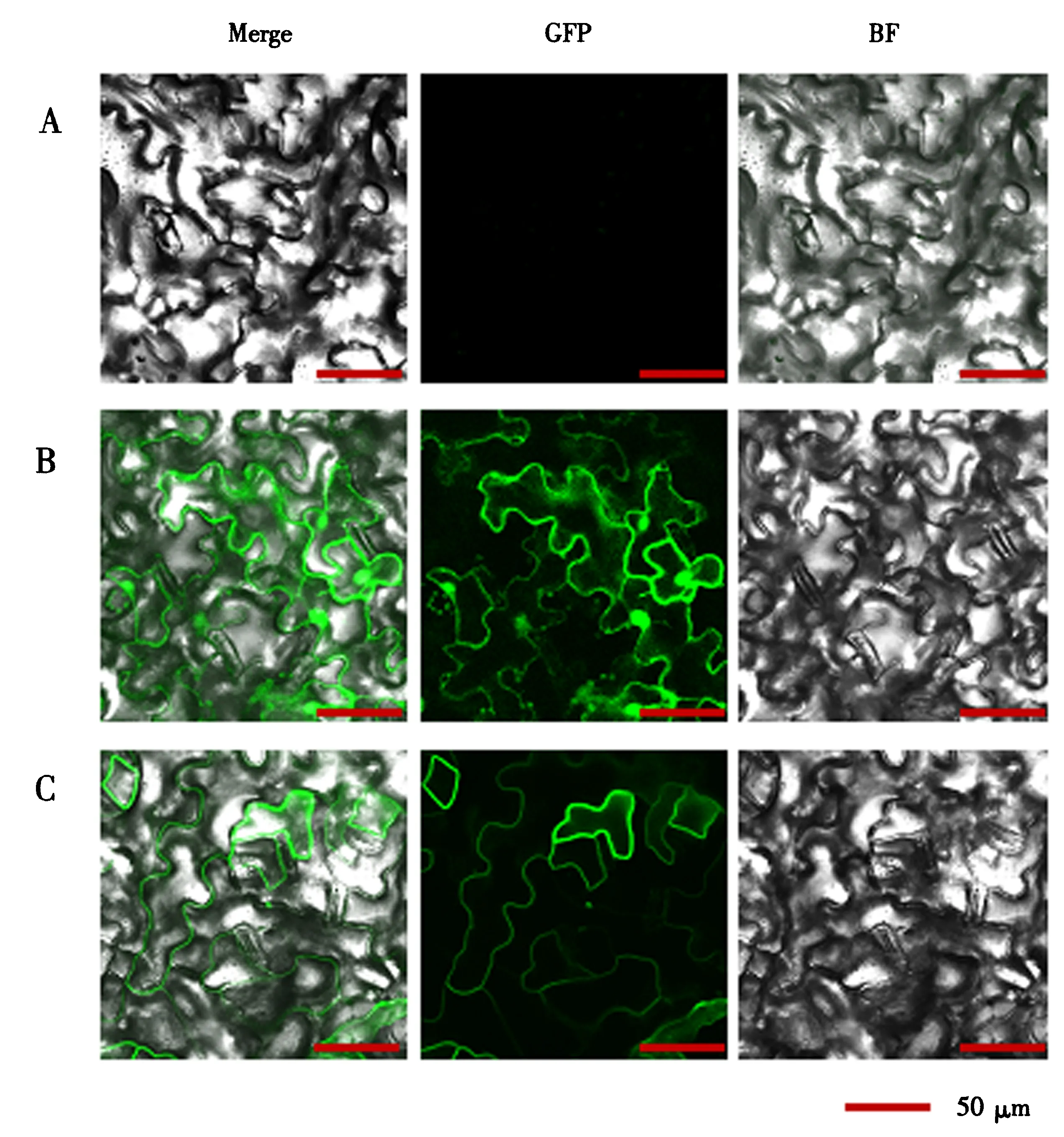
A.未经处理的油菜叶片;B.真空渗透表达GFP的油菜叶片;C.真空渗透表达YFP-BnSYP122的油菜叶片.BF.明场;GFP.绿色激发光场景;Merge.融合明场和激发光场景.图4 激光共聚焦验证GFP及YFP-BnSYP122在油菜叶片中的瞬时表达Fig.4 Transient expression of GFP and YFP-BnSYP122 in B.napus leaves verified by confocal laser
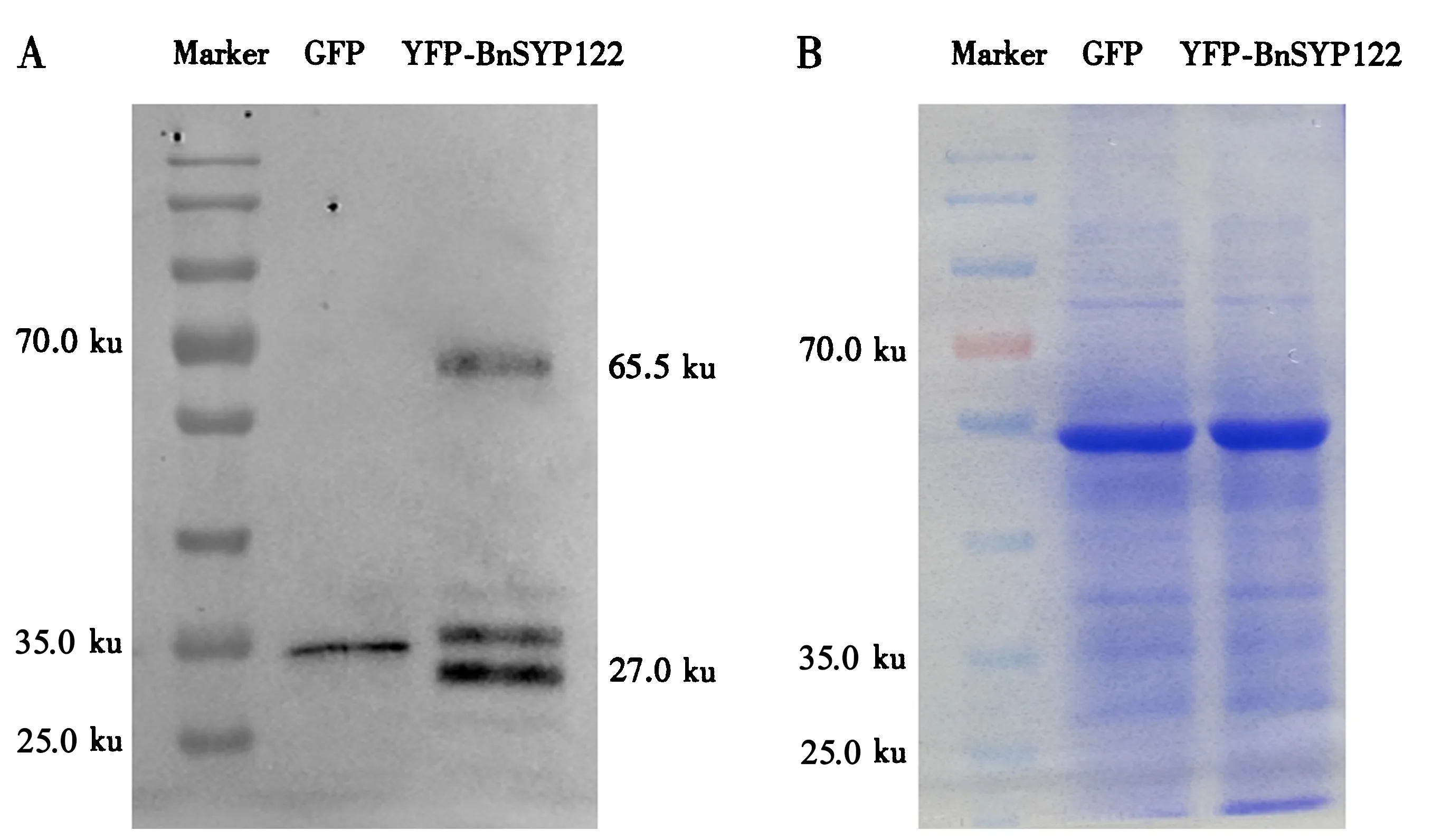
A.Western blot检测到的蛋白条带;B.考马斯亮蓝染色的总蛋白.图5 Western blot验证GFP和YFP-BnSYP122在油菜叶片中的表达Fig.5 Expression of GFP and YFP-BnSYP122 in B.napus leaves verified by Western blot
3 讨论
农杆菌介导的瞬时表达体系建立方法主要包括注射法和真空侵染法,2种方法在烟草中均能取得较高的表达效率.有报道称,由于油菜叶片特殊结构及叶脉的影响,农杆菌注射法无法在油菜叶片中取得较好的表达效果,而在子叶中可以成功完成瞬时表达[6].作者初期采用注射法在油菜叶片中表达GFP蛋白,效果也不理想.成熟植株的叶片具有更容易操作、表达蛋白获得量大、有利于后期功能研究等优势,因此,本研究测试真空侵染法是否能在油菜叶片中表达报告基因GFP和YFP融合的SNARE蛋白BnSYP122.
采用烟草的真空侵染体系,参考油菜的稳定表达方法,渗透缓冲液在MMA(10 mmol·L-1MgCl2,10 mmol·L-1MES,150 μmol·L-1乙酰丁香酮)的基础上,加入了Silwet L-77辅助侵染[19].采用真空侵染法在烟草叶片中表达外源蛋白,当压力为0.06 MPa时,只需要1~3 min就可以完成[20-21];而在油菜中双九号的叶片中,至少需要10 min.这说明农杆菌侵入油菜叶片比侵入烟草困难,因此,注射法无法取得较好的表达效果.真空侵染的最适时间与油菜苗龄和长势关系密切,4周龄的油菜叶片经真空侵染15~20 min可取得较好的效果,超过25 min可能导致叶片细胞坏死、萎蔫.农杆菌的菌液浓度与转化效率关系密切,浓度过高可能导致植物叶片萎蔫,过低则转化效率较低[6].本研究参考子叶瞬时表达设置农杆菌D600 nm为0.6~0.8,取得了较好的表达效果.通过农杆菌介导的真空侵染法将BnSYP122基因在油菜叶片中成功瞬时表达,并应用此方法成功表达了油菜的外源基因GFP.
据报道,SYP122定位于质膜[22],其所在亚家族Qa-SNARE共同的结构特征为一个N端的3个螺旋束(Ha、Hb、Hc)、一个由60~70个氨基酸组成的SNARE保守基序、一个Linker和位于C端的跨膜结构域[11].这与本试验结果高度一致,说明本研究建立的农杆菌真空侵染法不仅能够快速、方便地表达目的蛋白,还能够精准地观察目的蛋白的亚细胞定位.
猜你喜欢
当代水产(2022年1期)2022-04-26 14:35:30
中国现代医药杂志(2020年10期)2020-12-14 07:20:14
广州大学学报(自然科学版)(2019年1期)2019-05-07 01:33:26
现代检验医学杂志(2016年3期)2016-11-15 01:59:28
西南农业学报(2016年5期)2016-05-17 05:42:33
西南农业学报(2016年6期)2016-04-16 05:12:51
天津科技大学学报(2016年1期)2016-02-28 16:59:45
湖北师范大学学报(自然科学版)(2015年2期)2016-01-10 08:41:53
医学研究杂志(2015年3期)2015-06-10 06:41:52
特产研究(2015年1期)2015-04-12 06:36:20
