
罗汉果几丁质酶基因家族的生物信息学分析
2019-09-04 12:20李惠敏李璐璐陈玉梅李佳慧
生物信息学 2019年3期
李惠敏, 李璐璐, 陈玉梅, 李佳慧
(广西师范大学 生命科学学院,广西 桂林 541006)
罗汉果(Siraitiagrosvenorii)属葫芦科藤本落叶植物,具有一年种多年生的块根状宿根性,其药食两用的果实被誉为“东方神果”,全球90%以上的罗汉果产于广西桂林地区[1-2]。根结线虫病是一类危害大、分布广、防治难的植物病虫害,也是罗汉果生产中的主要病害之一。根结线虫不仅危害罗汉果植株根部,破坏根组织的分化和生理活动,导致地上部分生长发育迟缓、推迟开花结果或不开花结果,同时还会加重枯萎病、根腐病等土传性真菌病害和部分细菌病害的发生,有的果园未到结果盛期就因严重感染根结线虫病而被迫淘汰,损失极为严重[3-4]。因此,如何防治根结线虫病是目前罗汉果种植中迫于要解决的主要问题之一,而利用生物技术进行抗性育种被认为是在植物生物防治中最有前景的策略[5]。
几丁质酶( EC 3. 2. 1. 14)是一种能够将几丁质水解成N-乙酰葡萄糖胺的糖苷酶[6]。因真菌细胞壁均含有几丁质,线虫的卵壳和表皮中也含有几丁质物质,几丁质酶作为病原物和寄主植物相互作用中的一种重要蛋白质,已成为在植物抗病虫害研究中的热点[7]。植物虽然不含几丁质,但也存在着几丁质酶基因,植物几丁质酶可分布于植物的茎、叶、种子及愈伤组织中,分子量为25~55 kDa,多为单体,呈酸性或碱性,主要是内切几丁质酶,它随机水解几丁质内部β-1, 4- 糖苷键,释放出寡糖[8-12],植物几丁质酶部分家族成员还具有溶菌酶同源性[13-14],相关研究也表明植物几丁质酶的高水平表达能够增强植物对害虫、病原菌和环境胁迫的抵抗能力[10-12,15-16]。
随着高通量测序技术和生物信息学技术的发展,转录组测序技术(RNA sequencing,RNA-Seq) 目前被广泛应用于植物抗病性等相关基因的鉴定与分析[16-20],本研究基于RNA-Seq技术获得罗汉果根系转录组测序数据的基础上,对其中的几丁质酶基因家族成员进行生物信息学分析,期望为今后研究几丁质酶对罗汉果植株根结线虫病抗性分子机理提供参考,为罗汉果抗性育种提供基因来源。
1 材料与方法
1.1 材料
罗汉果为当地栽培品种青皮果,属于南方根结线虫易感品种,以罗汉果幼苗根系为植物材料进行转录组测序,委托上海派森诺生物科技有限公司Illumina HiSeq 2500 平台进行。
1.2 方法
1.2.1 罗汉果几丁质酶基因鉴定及理化特性分析
基于罗汉果幼苗根系的转录组测序数据及其注释结果,将搜索到的注释为几丁质酶的Unigene通过开放阅读框ORF Finder(https://www.ncbi.nlm.nih.gov/orffinder)和预测保守域CD Search(https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)在线软件检测确认,筛选出含GH18或GH19基因家族成员结构域特征且具有完整ORF的Unigene序列。将鉴定到的罗汉果几丁质酶基因编码的氨基酸序列使用ExPAsy (http//www.expasy.org/)进行在线预测其理化特性和跨膜结构域预测(http://www.cbs.dtu.dk/services/TMHMM)。
1.2.2 信号肽预测及亚细胞定位
利用TargetP[21](http://www.cbs.dtu.dk/services/TargetP/)和在线软件CELLO v2.5[22]对15条罗汉果几丁质酶序列进行信号肽预测和亚细胞定位。
1.2.3 罗汉果几丁质酶系统进化树构建及氨基酸序列结构分析
基于几丁质酶的氨基酸序列,采用MEGA6.0邻接法(Neighbor-Joining,NJ)缺失序列删除的方式,构建罗汉果几丁质酶系统进化树(bootstrap=1 000)。并以NCBI数据库(https://www.ncbi.nlm.nih.gov/)下载的19条拟南芥几丁质酶序列(本研究中将其编号AtCht1到AtCht19,序列相关信息见表1)为参考,对罗汉果几丁质酶进行分组。利用 DNAMAN软件、MEME和Phyre2[23]在线工具对罗汉果几丁质酶基因编码的氨基酸序列结构域进行分析。

表1拟南芥几丁质酶序列信息Table 1 Chitinases in Arabidopsis thaliana downloaded from NCBI
2 结果与分析
2.1 罗汉果几丁质酶基因家族成员的鉴定与理化性质分析
基于南方根结线虫侵染早期罗汉果根系转录组测序结果,搜索到注释为几丁质酶基因的Unigene共有28条,然后分别经ORF finder和CD-Search确认具有完整开放阅读框ORF和GH18或GH19基因家族成员结构域特征的Unigene,最后共鉴定到15条罗汉果几丁质酶Unigene,命名为SgCht1到SgCht15(见表2),ORF finder在线预测其编码的几丁质酶前体氨基酸数目最少的为SgCht6 (260个),最大的为SgCht12和SgCht15(均是343个)。利用 ExPASy在线软件对罗汉果几丁质酶基因编码的蛋白质进行理化性质预测结果见表2,罗汉果几丁质酶的分子量在27 kDa到37 kDa之间,多数为酸性蛋白质,只有SgCht1、SgCht4和SgCht9的理论等电点大于8.0;不稳定系数显示SgCht1、2、4、6、7、10、11和14为稳定蛋白,其余为不稳定蛋白;总平均亲水性显示SgCht1、2、9、12、13为疏水性蛋白,其余均属于亲水性蛋白。

表2 罗汉果几丁质酶特征Table 2 Characteristics of chitinases in Siraitia grosvenorii
2.2 信号肽预测及亚细胞定位结果
信号肽预测结果显示,本研究鉴定到的15个罗汉果几丁质酶前体其N端均有一段长度范围为19-30个氨基酸残基的信号肽(见表2),亚细胞定位结果显示这些几丁质酶均定位在胞外。
2.3 罗汉果几丁质酶系统发育分析及分类
植物中的几丁质酶主要为糖苷水解酶18家族和19家族成员(GH18和GH19),根据氨基酸序列特征,植物的内切几丁质酶分为5 类,GH18包含III类和V类植物几丁质酶, GH19包含Ⅰ、 Ⅱ、 Ⅳ类[13-14]。本研究以拟南芥几丁质酶的分类为参考,基于几丁质酶氨基酸序列的NJ系统发育树结果如图1所示,15条罗汉果几丁质酶序列(图中星号所示)归为3类即SgCht11属于I类;SgCht6、SgCht7和SgCht10属于IV类,其余为III类。
DNAMAN9多序列比对结果显示,Ⅰ类和IV类具有两个高度保守区,这两个保守区由一段富含脯氨酸、甘氨酸和精氨酸的可变交联区(见图2 ,虚线下划线所示)相隔开,N端高度保守区富含半胱氨酸(见图2,虚线框所示),其作用是与几丁质结合,简称CBD(Chitin-binding domain);C端是高度保守的催化区,有三个活性位点(见图2 实线方框所示)。IV类几丁质酶(SgCht6、SgCht7和SgCht10)与I类(SgCht11)相比有4 处缺失(见图2),氨基酸数目(见表1)明显比Ⅰ类少。
罗汉果GH18家族成员全部是III类几丁质酶,它们不含与几丁质结合的富半胱氨酸功能域, 而且仅有一个催化结构域,根据活性中心是否是谷氨酸残基分为几丁质酶和类几丁质酶(见图3,虚线框所示),SgCht1、2、4、8、9、12和SgCht15均有一个谷氨酸活性中心,SgCht5、SgCht13缺失了谷氨酸活性中心,而SgCht3和SgCht14的活性中性则分别是天冬氨酸和丙氨酸残基。
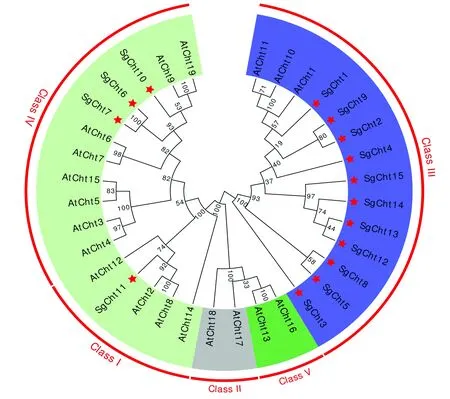
图1 罗汉果与拟南芥几丁质酶的系统发育树Fig. 1 Phylogenetic tree of chitinases from Siraitia grosvenorii and Arabidopsis thaliana

图2 GH19基因家族成员保守结构域氨基酸序列比对Fig.2 Alignment of conserved regions of GH19 amino acid sequences

图3 GH18 基因家族成员保守结构域氨基酸序列比对Fig.3 Alignment of conserved regions of GH18 amino acid sequences
2.4 罗汉果几丁质酶序列结构特点
利用 MEME 5.0.4软件对15个罗汉果几丁质酶的氨基酸序列进行motif查找,设置查找motif数目为6个,其余参数为默认值,结果如图4所示,GH18家族成员均具有motif1、motif2和motif4;而GH19家族成员仅有motif6。

图4 罗汉果几丁质酶蛋白基序Fig.4 Motifs of chitinase in Siraitia grosvenorii
GH18基因家族成员编码的蛋白质三级结构具有典型的(α/β)8桶状结构,即外部为 8 个α螺旋域,内部为 8 个平行的β折叠域[24],而GH19家族成员仅有α螺旋结构域。图5显示了罗汉果几丁质酶GH18基因家族成员SgCht12(见图5a)和GH19家族成员SgCht6(见图5b)由Phyer2软件预测的三级结构模型。其中,SgCht12 以c6idnA结构为模板,有 268个氨基酸残基与模板达到 100 %的可信度;SgCht6以c4mckA结构为模板,有199个氨基酸残基与模板达到100%的可信度。
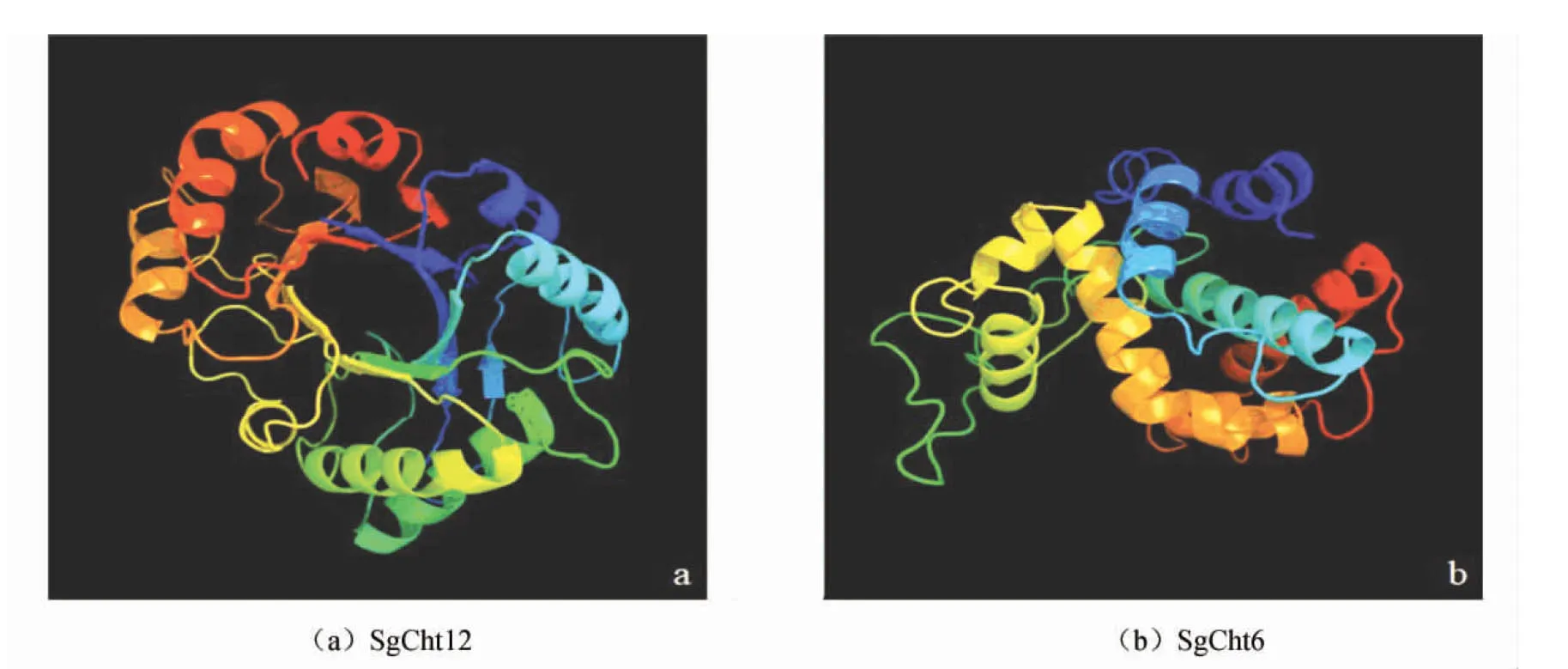
图5 三级结构模型
Fig.5 Tertiary structure of SgCht12(a)and SgCht6(b)
3 结论与讨论
罗汉果为广西特有经济植物,但罗汉果植株易感根结线虫病[3-4],被根结线虫感染后的罗汉果植株更容易被其他病菌的侵害,其中原因未见报道。几丁质是N-乙酰葡聚胺以β-1, 4-糖苷键连接起来的直链多聚物,是自然界中含量仅次于纤维素的一大类可再生聚合物,广泛存在于虾、蟹、昆虫、真菌等生物[25-26]。线虫卵壳及表皮的主要组成成分是几丁质,与真菌病原物一样,植物被线虫侵染时,可诱导产生几丁质酶[7]。几丁质酶作为病原物和寄主植物相互作用中的一种重要蛋白质,已成为在植物抗病虫害研究中的热点[5-7]。在植物抗病基因工程中,通过过量表达几丁质酶基因来提高植物抗病性相关研究已有报道[12,15,27-28]。郭悦等[12]将克隆到的花生几丁质酶基因AhChi进行遗传转化实验验证其生物学功能,结果表明转植株在黑斑病菌接种7d时其叶片褐化及坏死程度明显较非转基因植株要轻。谷天燕等[15]通过两端融合表达几丁质结合结构域的方法不但提高了几丁质酶对ɑ-几丁质和黑曲霉几丁质的结合能力和活性,同时也增强了其对病原真菌长枝木霉的抑制作用;南相日[27]的研究结果表明,转菜豆几丁质酶基因的马铃薯抗病性比对照提高了30%;王果萍等[28]也发现几丁质酶对镰刀菌引起的西瓜枯萎病有一定的抑制作用。随着对几丁质酶作用机理研究的深入[29-31],通过现代生物技术手段提高植物对真菌、昆虫、线虫等病害抗性将为有效生物防治提供新途径。本研究采用生物信息学方法从罗汉果根系转录组测序结果中鉴定到15个罗汉果几丁质酶基因,并对其编码的蛋白质进行了理化特性预测及系统发育分析,可为罗汉果几丁质酶基因生物学功能的鉴定提供理论基础,为罗汉果的抗病育种提供基因来源参考。本研究对转录组测序结果中的罗汉果几丁质酶基因的表达进行差异分析,结果发现接种根结线虫7 d, SgCht12基因和SgCht14出现显著性下调表达,分别是未接种根结线虫的0.17倍和0.21倍,这是否意味着已经被入侵的根结线虫建立了取食位点的罗汉果植株,很有可能由于几丁质酶的表达受到了抑制而导致对其他病菌的侵染更为敏感?这有待今后对罗汉果几丁质酶的生物学功能进行更深入的研究来进一步确定。
猜你喜欢
今日农业(2022年14期)2022-09-15
食品工业科技(2021年22期)2021-11-14
生物技术通报(2021年4期)2021-05-14
广东蚕业(2020年6期)2020-12-16
中成药(2018年11期)2018-11-24
中成药(2018年2期)2018-05-09
中国酿造(2017年8期)2017-09-03
浙江大学学报(农业与生命科学版)(2017年1期)2017-04-17
天然产物研究与开发(2016年6期)2016-06-05
食品与健康(2014年9期)2014-10-14
