
大豆蛋白-磷脂酰胆碱纳米乳液的显微观测及界面结构分析
2019-09-04 09:54吴长玲和铭钰史志玲王中江江连洲
中国食品学报 2019年8期
吴长玲 和铭钰 韩 瑞 史志玲 王中江 江连洲 李 杨,2*
(1 东北农业大学食品学院 哈尔滨150030 2 哈尔滨食品产业研究院 哈尔滨150030)
基于史梅尔散射光谱的理论预言,印度历史学家Raman 通过研究验证了散射光谱的存在[1],继而激光技术的发明推进了拉曼光谱技术的进步,使其成为研究生物分子结构的重要技术手段。而Fleischmann 通过对平滑银电极表面进行粗糙化处理吸附检测物,首次获得了吡啶分子的高分辨拉曼光谱[2],奠定了拉曼光谱在表面化学研究的实验基础。同时,激光共聚焦及多光子荧光显微成像已用于活细胞等生物化学样品的三维成像技术领域[3]。这与细胞成分的各种天然和人造荧光探针的发展相吻合[4]。对于不发荧光或不能耐受标记的化学物质或细胞成分,可以使用红外显微镜和自发拉曼显微镜作为基于振动特性的对比机制。常规的红外显微镜因使用的长波长而具有较低的空间分辨[5]。然而,本质上弱的拉曼信号需要高激光功率,并且通常被样品的荧光背景所掩盖。
显微观测技术在近20年以来得到突破性的发展,使分子表面科学进入一个更微观的世界。Shen 等[6]提出了基于相干反斯托克斯拉曼散射(CARS) 的多光子显微镜作为提供振动对比度的替代方法[7],然而,CARS 显微镜的敏感性受到非共振背景信号的限制,没有实现高分辨率的三维切片。纳米乳液乳滴界面组分复杂、形貌微小,窥测困难,乳液界面结构及微观形貌的分析手段较少。纳米乳液的研究需要更清晰的固/液、液/液原位的显微观测技术,如表面光谱及表面流变技术等。有关纳米乳液的界面吸附、结构舒张及构象重排研究,需对固/液及液/液界面进行切片以精细观测。随着现代化仪器的发展和显微观测技术的应用升级,以更微观和更清晰的纳米乳液乳滴界面观测技术解析纳米乳液界面组分分布及结构特征,逐渐成为现今此领域的研究热点。本研究拟采用激光显微共聚焦、拉曼光谱分析及拉曼成像技术,以观测分析超声及高压均质制备纳米乳液的界面结构及微观形貌特征,为纳米乳液制备及品质调控提供理论基础。
1 材料与方法
1.1 试验材料
大豆蛋白(蛋白含量89.21%),山东省高唐蓝山集团;大豆卵磷脂,上海楷洋生物技术有限公司;β-胡萝卜素,美国Sigma 公司;葵花籽油,中粮集团福临门压榨一级葵花籽油;试验基础试剂均为分析纯级,北京化学试剂公司。
1.2 试验仪器与设备
AL204 型分析天平,梅特勒-托利多仪器(上海)有限公司;PHS-3C 雷磁pH 计,上海精科;超声波细胞破碎仪,宁波新芝生物科技股份有限公司;布鲁克SENTERRA 激光拉曼光谱仪,杭州明凯科技公司;PerkinElmer Raman Station 400 拉曼光谱仪,美国PE 公司;A1 si 光谱型共聚焦,北京中晟天成科技有限公司;D-6L 超高压均质机,美国PhD 科技有限公司;Ultra-Turrax T25 高速分散器,德国IKA 公司;S22-2 型恒温磁力搅拌器,上海司乐仪器有限公司。
1.3 试验方法
1.3.1 粗乳液的制备 将一定量的大豆蛋白与磷脂酰胆碱混合溶解于98.5 mL 磷酸盐缓冲液中作为水相,另取0.01 g β-胡萝卜素溶于5 g 葵花籽油中作为油相,采用磁力搅拌器充分搅拌,直至β-胡萝卜素完全溶解。将油相与蛋白质水相溶液混合均一,用高速分散器14 000 r/min 分散5 min,获得粗乳液。
1.3.2 大豆蛋白-磷脂酰胆碱纳米乳液的超声制备工艺 分别取适量粗乳液于烧杯中,置超声波细胞破碎仪中,液面浸没变幅杆3 cm。制备条件:大豆蛋白添加量1.46%,大豆磷脂酰胆碱添加量0.21%,超声功率500 W,超声时间10 min,以循环冷热水控制超声温度。
1.3.3 大豆蛋白-磷脂酰胆碱纳米乳液高压均质乳化工艺 用高压均质机进行纳米乳液制备,制备条件:大豆蛋白添加量1.5%,大豆磷脂酰胆碱添加量0.22%,高压均质压力100 MPa,均质3 次,最终得到大豆蛋白-磷脂酰胆碱纳米乳液。
1.3.4 纳米乳液界面层组分结构特征的拉曼分析参考Herrero[8]方法。激发光波长532 nm,激光功率80 mW,扫描范围300~3 500 cm-1,每次扫描时间60 s,积分10 次,4 次扫描进行累加。以苯丙氨酸(1 003±1)cm-1作为归一化因子,绘制大豆蛋白及磷脂的拉曼谱图。谱图基线校正、谱峰归属查找采用ACD Labs V12 软件。
1.3.5 纳米乳液形态的激光共聚焦显微镜观测大豆蛋白-磷脂酰胆碱纳米乳液体系激光共聚焦显微镜的检测方法参考Puppo 等[9]的方法。大豆蛋白经尼罗蓝染液染色后呈现绿色荧光,葵花籽油经脂溶性荧光探针尼罗红染色后呈现红色荧光。分别将尼罗红(0.1%)和尼罗蓝(1%)溶解在丙醇中,漩涡混合30 s 后对大豆蛋白-磷脂酰胆碱纳米乳液染色30 min。染色结束后取10 μm 的乳液于载玻片上,采用激光共聚焦显微观测超声及高压均质制备大豆蛋白-磷脂酰胆碱纳米乳液的显微结构。
1.3.6 纳米乳液形态的拉曼显微镜观测 采用李雪等[10]及Yu T[11]等拉曼显微观测方法对超声及高压均质制备大豆蛋白-磷脂酰胆碱纳米乳液的界面结构进行分析。将待测液滴至于载玻片上,聚焦到液滴上,激光功率约5 mW,拉曼散射信号被聚焦到光谱仪(Spectra Pro 2300i,Acton,600 g/mm)的狭缝入口。根据扫描范围,设定x 轴和y 轴方向的扫描距离,启动程序打开 Win Spec 32 软件(光谱仪自带) ,收集扫描点的光谱。设定光斑每次移动步长为2 μm,并设定收集光谱积分时间,从而实现样品的全自动扫描。三维成像的部分是在二维成像的基础上,手动调节激光聚焦深度(即z 轴方向的步长) ,实现对样品的逐层扫描。各层的光谱收集方法与二维成像收集光谱方法相同。
1.4 数据处理
每组试验做3 次平行试验,对试验数据进行误差分析。采用统计学软件SPSS 18 对试验数据进行方差分析、相关性和差异显著性分析。采用Origin8.5 软件进行作图。
2 结果与讨论
2.1 纳米乳液形态的激光共聚焦显微镜观测
为了观测纳米乳液的微观形貌特征,采用激光共聚焦显微镜观察大豆蛋白-磷脂酰胆碱纳米乳液的显微结构,如图1所示。尼罗红所激发的荧光为红色,而纳米乳液自身发射的荧光极弱,可判断图1a 及1c 中红色荧光为纳米乳液内部油相中的尼罗红产生,油相呈现球状被较完整地包裹于乳滴内部。由于尼罗蓝侵染蛋白后所激发的荧光为绿色,因此判定图1b 及1d 中绿色荧光来源于乳液表面的大豆蛋白组分。由图1可知,大豆蛋白呈现球状形态,表明大豆蛋白完整地吸附于纳米乳液的界面处,呈核壳状结构。由于纳米乳液乳滴在水相中以布朗运动为主,在同一拍摄画面内,乳液乳滴出现“运动拖尾”现象,因此激光共聚焦显微镜观测粒径应大于实际粒径。亦因上述情况图中油相红色荧光标记图像并未与蛋白绿色荧光标记显微图像呈现完美重合。

图1 纳米乳液形态的激光共聚焦显微镜观测图Fig.1 Nano-emulsion morphology by laser confocal microscopy
2.2 纳米乳液界面层组分结构特征的拉曼分析
为了有效分析纳米乳液界面层组分结构特征,采用拉曼光谱技术进行分析探讨。由于蛋白质在水介质存在下荧光背景较强,所以难以对纳米乳液界面层组分结构进行有效分析。鉴于此,本研究借助布鲁克BRAVO 拉曼光谱仪具有良好的荧光消除特点,对纳米乳液界面分布的蛋白及磷脂结构特征进行分析,测试结果见图2。
大豆蛋白的构象主要由酰胺Ⅰ带的拉曼特征峰确定,酰胺Ⅰ带拉曼特征峰位置:α-螺旋结构:1 645~1 660 cm-1;β-折叠结构:1 665~1 680 cm-1;β-转角结构:1 680~1 690 cm-1;无规卷曲结构:1 660~1 670 cm-1。本试验中大豆蛋白的拉曼图谱二级结构的定量计算由Raman Spectral Analysis Package Version 2.1 软件完成。

图2 拉曼光谱图Fig.2 Raman spectra

表1 拉曼光谱峰位归属Table 1 Raman spectroscopy peak attribution

表2 不同方式制备乳液中蛋白二级结构组分含量Table 2 Percentages of protein secondary structure of sample with different treatment
由上表中蛋白质二级结构组成可知,两种方式制备的纳米乳液界面处大豆蛋白的二级结构中均以α-螺旋结构及无规卷曲结构为主要构象单元,β-折叠及转角结构含量均较低,验证了在谱图中1 655 cm-1处出现明显α-螺旋结构——拉曼归属峰。由此推测在大豆蛋白-磷脂酰胆碱纳米乳液形成的吸附重排作用下,大豆蛋白结构趋向于α-螺旋及无规卷曲的二级结构组成。研究表明[12],蛋白在界面吸附过程中更趋向于有序的二级结构,验证了本研究中α-螺旋结构含量增加。
比较可知,高压均质制备的纳米乳液界面处蛋白无序二级结构单元含量较高,而有序结构单元组成含量较低。研究发现β-折叠结构含量增加与蛋白质间交互作用有关[13]。另有研究指出O/W乳化界面处蛋白质交互作用下分子间β-折叠结构含量增加[14],而在超声处理下大豆蛋白表现出显著的蛋白聚集作用,在本研究中β-折叠结构含量增大。
色氨酸在拉曼谱图上表现出多个拉曼谱带用于监测蛋白质微环境极性及氢键变化规律。Li-Chan[15]研究表明760 cm-1附近区域的拉曼峰强度降低与色氨酸残基由原本“包埋式”转变为“暴露式”有关。已有研究表明热变性造成蛋白质结构的破坏,进而引起色氨酸残基的暴露,在拉曼谱图中表现为色氨酸谱带强度的降低[16-19]。比较可知,在本研究中,超声制备的纳米乳液中大豆蛋白在760 cm-1附近区域的色氨酸拉曼峰强度较低,该区域归属于色氨酸残基的伸缩振动,表明此条件下色氨酸残基更趋于“暴露态”。

表3 不同处理条件下大豆蛋白侧链基团谱带强度Table 3 Intensities of tryptophan band,tyrosyl doublet of soy protein at different treatment
850 cm-1和830 cm-1是酪氨酸残基苯环的呼吸振动和面外弯曲倍频之间的费米共振[19]。通过两条谱线的强度比,推测酪氨酸是氢键的供体或是受体。若I850/I830比值为2.5,则酪氨酸苯环上的羟基氧原子是强氢键受体;若I850/I830比值为1.25,则酪氨酸苯环上的羟基氧原子为中强度氢键的供体或受体;若I850/I830比值为0.3,表明酪氨酸的苯环上的羟基氧原子是强氢键的供体。本研究中,I850/I830比值分布于0.98~1.11,表明所测试蛋白的酪氨酸残基暴露于溶液的极性微环境下,作为中性强度氢键的供体或受体。
通过比较可知,超声制备纳米乳液界面蛋白的酪氨酸拉曼归属峰强度I850/I830比值高于高压均质制备的,由此推测超声制备纳米乳液界面蛋白的酪氨酸更趋于“暴露态”。综合上述,超声制备纳米乳液界面蛋白的疏水性基团暴露程度高于高压均质,疏水性基团的“暴露”将增加蛋白与油相的分子间作用,增强乳化稳定性。
1 450 cm-1是脂肪族氨基酸的拉曼归属谱线,研究表明脂肪族氨基酸的拉曼归属峰强度降低与脂肪族氨基酸暴露有关[20]。本研究中,超声处理及高压均质作用下大豆蛋白在1 450 cm-1处的拉曼归属峰均较高,脂肪族氨基酸趋于“包埋态”,这与大豆蛋白在界面吸附过程中的结构重排有关。另外,脂肪族氨基酸与油相组分的疏水交互作用也是脂肪族氨基酸“内埋”的可能原因。比较可知,高压均质制备纳米乳液界面蛋白的脂肪族氨基酸拉曼归属峰值较大,表明此条件下脂肪族氨基酸更趋于“包埋态”。
拉曼光谱中的C-C 骨架振动可以用来表征磷脂酰胆碱脂链的反式-旁式构象变化。面内和面外的C-C 伸缩振动出现在1 000~1 200 cm-1区域,其中1 064 cm-1和1 129 cm-1谱线代表C-C 链反式构象的伸缩振动,而1 090 cm-1谱线归属于C-C 键扭曲式异构体的贡献[21-22]。本研究中考虑到大豆蛋白的拉曼吸收,采用差谱对磷脂酰胆碱的结构进行光谱分析,通过I1090/I1064及I1090/I1129表示脂链的无序程度,具体结果见表4。

表4 不同处理条件下磷脂酰胆碱I1090/I1064 及I1090/I1129 拉曼峰强度Table 4 Normalized intensities of the I1090/I1064 and I1090/I1129 SPI-Lec with different treatment
由表4可知,高压均质制备纳米乳液界面处磷脂酰胆碱的I1090/I1064及I1090/I1129拉曼峰强度较高,表明此制备方式下磷脂酰胆碱脂链中C-C 的旁式构象更多,脂链更为无序。由此可推断磷脂酰胆碱在高压均质方式制备纳米乳液的界面处表现出与大豆蛋白更强的疏水交互作用。
2 880和2 850 cm-1谱线分别属于CH2基团的C-H 对称伸缩振动和反对称伸缩振动模式,I2848/I2880可表征脂链侧向耦合能力以及有序-无序排列[23]。对乳铁蛋白及其肽段与磷脂间的交互作用的研究表明,磷脂的拉曼归属峰I2880/I2850比值较低,且乳铁蛋白及其肽段与磷脂间交互作用下两者比值变化率较低,推断此条件下乳铁蛋白及其肽段有可能仅在脂质体表面与磷脂分子的极性头部基团发生作用,而未深入到脂质体的疏水内核与脂酰链相互作用,即未引起磷脂脂酰链结构的改变。对比已有研究,本研究中两种方式制备的纳米乳液界面磷脂酰胆碱脂链的拉曼归属峰I2848/I2880比值较高,侧链间无序性较强,推测磷脂酰胆碱已深入油相内部或与界面蛋白间发生显著的疏水作用。比较可知,高压均质制备纳米乳液界面处磷脂酰胆碱的拉曼归属峰I2848/I2880比值更高,表明此条件下磷脂酰胆碱的尾部疏水性结构更加无序,磷脂酰胆碱与油相或界面蛋白间发生显著的疏水作用,进而稳定纳米乳液体系。
2.3 纳米乳液形态的拉曼显微镜观测结果
生物物质分子中含有许多特殊的化学键或者官能团,这些化学键或官能团的存在与否,可用于判断该区域是否含有这种物质,这种物质的多少也可由这种化学键在单位面积内的含量表示。特征峰的峰面积或峰高可以反映物质含量。在选定的组织区域进行逐点扫描,收集各个微区的拉曼光谱,抽取特征峰计算峰面积,根据归一化结果即可实现拉曼成像观测。

图3 拉曼成像图Fig.3 Raman image
为了观测纳米乳液界面乳化层结构,利用拉曼显微成像技术对纳米乳液的乳滴进行观测。由于大豆蛋白在1 660 cm-1处具有显著的拉曼吸收,红框区内选定单一纳米乳液乳滴进行分析,随着观测位移逐渐探向油相,形成6 张乳滴拉曼成像图。由图4中U-1 及H-1 曲线可知,在乳滴界面处分布大量绿色标示的大豆蛋白组分,通过拉曼成像色差及拉曼强度对比可知,超声制备的纳米乳液乳滴界面处分布更多的大豆蛋白组分。随着观测视角逐渐探向油相,界面处绿色的大豆蛋白分布区域逐渐缩小,展现出更多的蓝色油相区域,绿色大豆蛋白更多地分布于乳滴界面处。由此可知,大豆蛋白应均匀地分布于乳滴界面。
分析观测区域的拉曼光谱可知,1 665 cm-1处表现出大豆蛋白酰胺I 带的特征归属峰,而在2 840 及2 880 cm-1处主要是磷脂酰胆碱的CH2基团的C-H 对称伸缩振动和反对称伸缩振动归属峰。由此表明乳滴界面处分布了大豆蛋白及磷脂酰胆碱。随着观测视角逐渐探向油相,整体拉曼光谱的强度有所降低,这可能与大豆蛋白在观测区域分布减少有关,而此过程中大豆蛋白及磷脂酰胆碱的特征峰并未消失,表明乳滴界面处大豆蛋白与磷脂酰胆碱均匀分布及可能的交互作用。
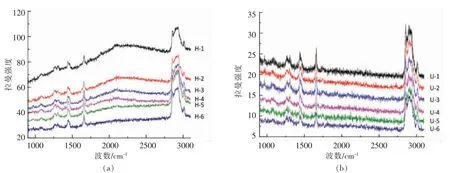
图4 拉曼光谱图Fig.4 Raman spectra
3 结论
采用激光显微共聚焦观测超声及高压均质制备大豆蛋白-磷脂酰胆碱纳米乳液的显微结构,研究发现大豆蛋白呈现球状形态,表明大豆蛋白完整地吸附于纳米乳液的界面处,呈核壳状结构。借助布鲁克BRAVO 拉曼光谱仪具有良好的荧光消除特点,对纳米乳液界面分布的蛋白及磷脂结构特征进行解析,研究发现大豆蛋白-磷脂酰胆碱纳米乳液形成中,界面吸附重排作用下,大豆蛋白结构趋向于α-螺旋及无规卷曲的二级结构组成。超声制备纳米乳液的界面蛋白疏水性基团暴露程度高于高压均质,疏水性基团的“暴露”增加了大豆蛋白与油相的分子间作用,增强乳化稳定性,磷脂酰胆碱在高压均质制备的纳米乳液的界面处表现出与大豆蛋白更强的疏水交互作用。
猜你喜欢
现代食品(2022年11期)2022-07-18
化工管理(2021年27期)2021-10-20
辽宁省博物馆馆刊(2021年0期)2021-07-23
上海理工大学学报(2021年3期)2021-07-20
物理学报(2020年16期)2020-08-29
当代水产(2020年4期)2020-06-16
保健与生活(2019年22期)2019-11-25
新疆人文地理(2016年9期)2016-05-14
癌变·畸变·突变(2015年3期)2015-02-27
汽车与新动力(2013年1期)2013-03-11
