
过表达SHC3激活AKT/Nrf2/GSH通路保护帕金森病氧化应激损伤
2019-09-04 09:14齐献忠邢英瀛秦慧兵
中国老年学杂志 2019年17期
齐献忠 邢英瀛 秦慧兵
(南阳市中心医院神经内科,河南 南阳 473000)
帕金森病(PD)主要是由中枢神经系统脑细胞中多巴胺能神经元的损失造成的,通常表现为运动障碍〔1,2〕。PD在神经退行性疾病发病率仅次于阿尔茨海默病,其发病率约为全球65岁以上人口的2%〔3〕。慢性活性氧(ROS)水平的增加或ROS消除能力的降低均称为氧化应激(OS)〔4〕。研究显示,OS在多巴胺神经元的退化、死亡中发挥关键性作用〔5〕。目前,尚未发现有效的可保护或逆转PD的神经退行药物。含有Src同源结构域2的衔接蛋白(SHC)家族包括SHCA、SHCB、SHCC(SHC3)、SHCD,其在哺乳动物中均具有特异的原型信号传导细胞,其中SHC3已被证明仅限于神经元细胞〔6〕且在典型成熟神经元中表达〔7〕。据报道,磷酸肌酶-3-激酶(PI3K)-蛋白激酶B(AKT)-FoxO、PI3K-AKT、PKCβ/p66Shc、FoxO/Wnt、p66Shc/IQGAP1/核因子NF-E2相关因子(Nrf)2及AKT/Nrf2/谷胱甘肽(GSH)等通路均参与调控OS〔8~11〕。其中PI3K-AKT-FoxO和AKT/Nrf2/GSH通路调控PD中的氧化应激已得到证实〔11〕。但SHC3与AKT/Nrf2/GSH通路在PD中的作用关系尚未清楚。本研究拟分析过表达SHC3对PC12细胞氧化应激、凋亡及细胞活性的影响。
1 材料与方法
1.1材料 PC12细胞购自上海生命科学研究院细胞中心;1-甲基-4-苯基吡啶离子(MPP+)购自瑞士SIGMA;乳酸脱氢酶(LDH)、ROS、丙二醛(MDA)、GSH测定试剂盒购自赛默飞公司;聚氰基丙烯酸正丁酯(BCA)蛋白定量试剂盒、qRT-PCR试剂盒、噻唑蓝(MTT)、脂质体LipofectamineTM2000、逆转录试剂盒购于大连Takara公司;聚偏氟乙烯(PVDF)膜、十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(SDS-PAGE)试剂盒、电化学发光(ECL)液和RIPA蛋白裂解液均购于碧云天生物技术公司;Nano-Drop 2000微量分光光度计(美国Thermo公司);ABI 7500型实时荧光定量PCR系统购于美国ABI公司;细胞培养箱购于美国Forma Scientific公司;PCR 仪购于美国BIO-RAD公司。
1.2细胞培养及分组 用含10%胎牛血清的DMEM培养基培养PC12细胞,置于5% CO2的37℃培养箱中常规培养。正常组:取上述培养细胞不做任何处理;模型组:用250 mmol/L的MPP+处理PC12细胞24 h,构建PD细胞模型;模型+NC组:将载体pcDNA3.1用脂质体法转染至PC12细胞,转染成功后,按照模型组操作处理细胞;模型+SHC3组:将pcDNA3.1-SHC3转染至PC12细胞,转染成功后,按照模型组操作处理细胞。将以上各处理组细胞进行qRT-PCR实验、Western印迹实验、酶联免疫吸附试验(ELISA)。
1.3qRT-PCR实验 取适量对数生长期各组细胞进行Trizol法提取细胞总RNA,并用Nano-Drop 2000微量分光光度计进行RNA定量。DNaseⅠ消化RNA中可能污染的DNA。逆转录反应采用逆转录试剂盒方式,操作按照试剂盒说明书进行,合成模板链cDNA。按照反应体系进行,每个样品重复3次,取平均值,反应结束后通过分析Ct值,计算定量结果,以2-△△Ct法测定SHC3 mRNA的相对表达水平。
1.4Western印迹实验 取适量对数生长期各组细胞,用RIPA蛋白裂解液进行冰上蛋白裂解1 h,提取总蛋白,以BCA法测定样品蛋白的浓度。以4∶1的比例加入蛋白上样缓冲液(×5),混匀,沸水浴变性5 min,离心取上清,取60 μg目的蛋白、蛋白Marker进行SDS-PAGE。然后将蛋白湿转至PVDF膜,5%脱脂奶粉室温封闭1 h,TBST洗涤,4℃条件下,将封闭后的PVDF膜放入1∶1 000倍稀释的一抗中反应过夜,以封闭液洗膜3次,每次5 min,再在37℃下将PVDF膜转入1∶2 000倍稀释的二抗中反应2 h。于暗室内ECL试剂盒显影曝光:滴加化学发光剂显影,以凝胶成像系统采集图像。Quantity One 4.62图像分析软件进行条带灰度分析,以β-actin为内参,以目的条带灰度值与β-actin灰度值的比值表示目的蛋白SHC3、B细胞淋巴瘤-2基因(Bcl)-2、Bcl-2关联X蛋白(Bax)、含半胱氨酸的天冬氨酸蛋白水解酶(caspase)-3、p-AKT、Nrf2的表达情况。
1.5ELISA试验 取适量对数生长期各组细胞,离心取上清。
1.6MTT实验 取适量对数生长期各组细胞,加入20 μl、5 g/L的MTT溶液,继续培养4 h,取出后吸去上清,每孔加入150 μl二甲基亚砜(DMSO),震荡待结晶充分溶解,在490 nm波长下检测细胞吸光度(A)。每组设5个重复孔,实验重复3次。细胞活性与A490值呈正相关。
1.7膜联蛋白(Annexin)V-FITC/碘化丙啶(PI)细胞凋亡检测 取适量对数生长期各组细胞,用预冷磷酸盐缓冲液(PBS)洗涤 2次。按照Annexin V-FITC/PI细胞凋亡检测试剂盒中的说明书要求操作,最后用流式细胞仪检测并分析测定结果。每个样品重复3次。细胞凋亡率(%)=早期凋亡率(Annexin V+/PI-)+晚期凋亡率 (Annexin V+/PI+)。
1.8统计学处理 采用GraphPad Prism7.0进行单因素方差分析、SNK-q检验。
2 结 果
2.1构建过表达SHC3的PC12细胞模型 与正常组相比,模型组、模型+NC组SHC3 mRNA和SHC3蛋白表达量均显著降低(P<0.05),模型+SHC3组SHC3 mRNA和SHC3蛋白表达显著升高(P<0.05);与模型+NC组相比,模型+SHC3组SHC3 mRNA和SHC3蛋白表达显著升高(P<0.05),见表1,图1。
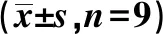
表1 各组细胞中SHC3 mRNA、SHC3蛋白表达量
与正常组比较:1)P<0.05;与模型+NC组比较:2)P<0.05;下表同
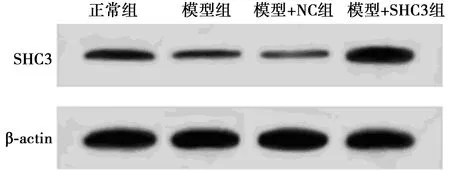
图1 各组SHC3的蛋白表达
2.2过表达SHC3对MPP+诱导PC12细胞氧化应激损伤的影响 与正常组相比,模型组、模型+NC组、模型+SHC3组ROS、MDA含量显著升高,GSH含量显著降低(P<0.05);与模型+NC组相比,模型+SHC3组ROS、MDA含量显著降低,GSH含量显著升高(P<0.05),见表2。

表2 各组氧化应激分子ROS、MDA、GSH的含量
2.3过表达SHC3对MPP+诱导PC12细胞活性的影响 与正常组相比,模型组、模型+NC组、模型+SHC3组LDH含量显著升高,细胞活性显著降低(P<0.05);与模型+NC组相比,模型+SHC3组LDH含量显著降低,细胞活性显著升高(P<0.05),见表3。
表3 各组LDH含量和细胞活性比较
2.4过表达SHC3对MPP+诱导PC12细胞凋亡的影响 与正常组相比,模型组、模型+NC组、模型+SHC3组细胞凋亡率、Bcl-2/Bax蛋白表达量、caspase-3蛋白表达量显著升高(P<0.05);与模型+NC组相比,模型+SHC3组均显著降低(P<0.05),见表4,图2。
2.5过表达SHC3激活AKT/Nrf2/GSH通路 模型组、模型+NC组、模型+SHC3组与正常组p-AKT、Nrf2蛋白表达量差异有统计学意义(P<0.05);模型+SHC3组与模型+NC组相比,p-AKT、Nrf2蛋白表达量显著升高,见表5,图3。
表4 各组细胞凋亡率及凋亡蛋白Bcl-2/Bax、caspase-3表达量比较

图2 各组细胞中Bcl-2、Bax、caspase-3蛋白表达

组别p-AKTNrf2正常组1.00±0.051.00±0.06模型组0.72±0.091)0.63±0.071)模型+NC组0.79±0.091)0.59±0.061)模型+SHC3组1.84±0.171)2)1.66±0.161)2)F值67.01578.196P值0.0000.000

图3 各组p-AKT、Nrf2蛋白表达
3 讨 论
PD作为世界第二常见的中枢神经系统退行性疾病,在全球的患者超过10万例〔12,13〕。虽然PD在动物模型中进行了广泛的研究,但仍缺乏有效的保护PD神经药物〔14〕。SHC蛋白家族的特征是(CH2)-PTB-CH1-SH2模块化。最初从果蝇中的一个位点(dShc)演变到哺乳动物中的3个位点(shc,rai和sli)。由于其启动密码子的剪接,哺乳动物的3个基因至少编码6种Shc蛋白,且这些蛋白质均具有与氨基末端的磷酸酪氨酸激酶受体结合的PTB结构域和CH2结构域及与Grb2结合的CH1结构域。SHC3蛋白由两种异构体(p66SHC3和p52SHC3)组成,前者比后者多了一个CH2结构域。SHC3仅在成熟神经细胞中表达〔15〕。Troglio等〔16〕发现,Rai可提高小鼠神经元细胞的缺氧或氧化应激诱导的细胞凋亡并增加缺血再灌注诱导的小鼠神经细胞凋亡和梗死面积,其机制可能为Rai作为应激反应基因,在低氧或氧化损伤后激活PI3K/AKT,揭示Rai在脑损伤中具有功能性神经保护作用。Gong等〔17〕研究表明,沉默SHC3可导致运动迟缓,且沉默SHC3抑制AKT和FoxO磷酸化的程度,失活PI3K/AKT/FoxO信号通路。本研究揭示了过表达SHC3可抑制PC12细胞凋亡。
氧化应激是机体或细胞内氧自由基的产生与清除失衡,导致ROS和活性氮在体内或细胞内蓄积而引起的氧化损伤过程,其常见的检测指标有ROS、SOD、MDA、一氧化氮(NO)和GSH〔18〕。GSH参与H2O2的还原反应,其为脑内最重要的自由基清除系统。MDA为过氧化脂质的降解产物,其含量可反映过氧化脂质的水平。ROS为分子氧经过线粒体代谢的副产物,其含量可反映氧化应激强度。LDH为细胞质酶,在细胞膜通透性增高时,LDH露出细胞外,反映细胞的损伤及活力〔19〕。孟凛冽等〔20〕研究表明,黑质多巴胺神经元缺损与线粒体功能障碍、钙超载、铁离子堆积、免疫炎症等造成的氧化性损伤有关,且其可促进多巴胺能神经元凋亡。本研究发现,过表达SHC3可保护MMP+诱导的PC12细胞氧化应激水平,增强细胞活力。
Nrf2是维护氧化还原平衡的关键调节因子,尤其是在持续激活PI3K-AKT信号的条件下。活化的PI3K-AKT通路增强Nrf2在核内的积累,使Nrf2促进具有抗氧化、抗炎、抗凋亡基因的表达〔21,22〕。Du等〔23〕研究发现,人参主要活性成分(Rg1)对铁毒性神经具保护作用,其机制为激活AKT/Nrf2途径及增加Nrf2诱导的HO-1和Cu/Zn SOD表达的抗氧化,为PD发病机制及治疗提供依据。Gunjima等〔24〕研究发现,3,4-二羟基苯甲酰丙酮(DBL)可逆转SH-SY5Y细胞的PD相关神经毒素6-羟基多巴胺的神经毒性,其机制为通过PI3K/AKT激活Nrf2/GSH途径。本研究发现,过表达SHC3可逆转MMP+诱导损伤PC12细胞中p-AKT、Nrf2蛋白表达及GSH含量,揭示过表达SHC3激活AKT/Nrf2/GSH通路。 综上,过表达SHC3可保护PD的氧化应激损伤,其机制与激活AKT/Nrf2/GSH通路有关,为PD的治疗及发病机制的研究奠定理论基础。
猜你喜欢
湖北农业科学(2022年11期)2022-07-18
昆明医科大学学报(2022年4期)2022-05-23
上海交通大学学报(医学版)(2022年3期)2022-05-05
河北农业大学学报(2022年2期)2022-04-26
电子产品世界(2021年8期)2021-01-16
实用肿瘤学杂志(2020年4期)2020-12-08
世界科学技术-中医药现代化(2020年2期)2020-07-25
中国疼痛医学杂志(2019年9期)2019-01-04
创新时代(2016年8期)2016-10-21
弹箭与制导学报(2015年1期)2015-03-11
