
江淮地区不同轮茬作物对苜蓿产量及根际土壤质量的影响
2019-08-30 02:26:42李争艳徐智明师尚礼贺春贵
草业学报 2019年8期
李争艳,徐智明,师尚礼,贺春贵
(1.甘肃农业大学草业学院,甘肃 兰州 730070; 2.安徽省农业科学院畜牧兽医研究所,安徽 合肥 230031)
紫花苜蓿(Medicagosativa)是世界广泛种植的优质牧草,它具有一种特殊的种内化感作用—自毒性[1],苜蓿生长多年会导致土壤自毒物质的积累,而且随着苜蓿生产年限的增加,土壤养分及水分会显著下降,导致土壤环境恶化,使苜蓿产量品质下降[2]。随着我国奶业不断发展,对优质牧草的需求不断加强,国内苜蓿种植面积增加、范围扩大,逐步形成产业化及规模化态势,与此同时,土传病害、自毒效应及连作问题也日益严重[3]。科学轮作是解决连作障碍最有效的措施[4],然而,对于江淮地区的苜蓿生产,缺少相关经验,且高温高湿的环境更容易引发病虫害及连作障碍,对该地区苜蓿生产造成严重的制约和影响。因此,本试验通过研究苜蓿长期生长及不同轮作方式下土壤物理化学及生物学性质,来研究不同轮茬作物对土壤质量、地力及苜蓿产量的影响,为江淮地区苜蓿种植方式的建立提供理论依据。
来自植物根、土壤微生物、植物和动物残留物的土壤酶是土壤肥力、有机质分解、土壤生物活性和代谢速度的良好指示物[5],土壤酶活性的降低被认为是导致连作障碍的主要因素[6]。土壤微生物群落结构受到作物连作、轮作[7]、施肥制度[8]、灌溉[9]等农业措施的影响,也是土壤健康的重要决定因素。微生物群落变化可导致土壤代谢能力和土壤质量的变化。同时,土壤微生物多样性在可持续农业中发挥着重要作用,微生物可以通过影响其他的生化过程诸如植物养分循环、土壤物理结构维持及化学降解而促进农业生产。连作能够引起根际土壤中微生物群落的变化[10],导致微生物种群结构失衡,有害的微生物会对植物生长产生负面影响,形成连作障碍[11]。国内外专家已经有很多有关轮作缓解连作障碍的研究,对黄瓜(Cucumissativus)、马铃薯(Solanumtuberosum)、小麦(Triticumaestivum)、棉花(Gossypiumhirsutum)、高粱(Sorghumbicolor)等作物的研究表明,与单一连作的种植模式相比,在相同管理条件下与不同作物轮作能改变土壤微生物组成,缓解连作障碍,进而增加作物产量,提高作物品质[6-7,10,12-13]。张立成等[14]研究表明,稻(Oryzasativa)-稻-油菜(Brassicachinensis)轮作增加了细菌群落的丰度。陈丹梅等[15]研究表明,轮作模式通过影响烟草(Nicotianatabacum)的土壤酶活性及真菌群落,进而影响产量。李戌清等[16]研究表明,轮作较连作有利于提高土壤酶活性,增加细菌多样性,保持地力。鉴于江淮地区的牧草种植模式科学有效办法的缺乏,研究了苜蓿长期生长及轮茬高粱(Sorghumbicolor)和玉米(Zeamays)对苜蓿产量、土壤养分、土壤容重、酶活性及根际土壤细菌群落多样性的影响,以期为江淮地区牧草生产经营管理提供技术支撑,为草地生态环境保护提供理论指导。
1 材料与方法
1.1 材料和样地
试验在安徽省五河县朱顶镇(117°26′-118°04′ E, 32°55′-33°20′ N)秋实草业公司试验基地进行。海拔16 m,年降水量800~1000 mm,年平均气温15.1 ℃,无霜期216 d,土壤类型为典型的黄潮土。设3个处理:连续生长4年苜蓿灭茬重新种植1年为对照(CK),连作3年苜蓿-玉米-苜蓿(T1),连作3年苜蓿-高粱-苜蓿(T2),试验小区13 m×6 m,每个小区分为3个重复。苜蓿供试品种为Magna601,由克劳沃(北京)生态科技有限公司提供。苜蓿种植密度为60万株·hm-2,每年刈割5次,留茬高度5 cm。玉米供试品种为先玉047,高粱供试品种为海牛(光敏型,Monster),由百绿(天津)国际草业有限公司提供。玉米行距40 cm,穴播种植,密度9万株·hm-2;高粱行距35 cm,穴播种植,密度12万株·hm-2。2012-2015年连续4年(CK)苜蓿地施肥水平均为(N, 180 kg·hm-2; P2O5, 260 kg·hm-2; K2O, 320 kg·hm-2);2015年4月轮茬处理(T1、T2)地块灭茬后,深耕翻地,玉米与高粱同时播种,施肥水平均为(N, 200 kg·hm-2; P2O5, 240 kg·hm-2; K2O, 300 kg·hm-2),苜蓿刈割5次,留茬高度5 cm,高粱刈割2次,分别于2015年7月20日和2015年9月20日刈割,留茬高度10 cm,玉米刈割1次,于2015年9月20日刈割,重复3次;2016年4月所有处理统一种植苜蓿,种植方案与前4年苜蓿种植方案一致。整个试验期均不灌溉,苗期人工除草1次,2016年10月采集全部土壤样品。播种前0~20 cm土层土壤pH值7.49,有机质19.27 g·kg-1,总氮1.53 g·kg-1,有效磷8.37 mg·kg-1,速效钾167.77 mg·kg-1。
1.2 苜蓿产量测定
2016年从5月上旬开始到10月上旬,共刈割5次,测定鲜草产量及其水分,计算干物质产量(采用105 ℃烘干法测定)。产量为单位面积土地上所收获地上部分的全部产量,每个小区测产面积5 m2,重复3次,留茬高度5 cm。
1.3 土壤测定项目及采样方法
于2016年10月,用直径2.5 cm土钻按S点取样法在每个样地选5个点土壤样品,深度为0~20 cm。在清除表面覆盖物后取样,将5个样品混为1个土样,放入无菌自封袋。每个样品分3份,1份保存在-80 ℃进行DNA提取,1份风干,并过2 mm筛进行化学分析,1份鲜样送达试验室立即进行酶活性检测。另外在距离植株15 cm处挖剖面分层取样,土层深度分别为0~10 cm,10~20 cm和20~30 cm,同一层不同位置用环刀取3个重复样本,用铝盒现场称湿重并放入自封袋,带回实验室测土壤容重。
1.4 土壤理化性质及土壤酶活性测定
通过在去离子水与土壤按1.0∶2.5体积比混合的悬浮液中插入玻璃电极计来测量土壤pH值。采用环刀法测量土壤容重,采用重铬酸钾法对有机质含量进行定量,总氮和有效磷分别用凯氏定氮法和钼锑抗分光光度法进行定量,速效钾用火焰光度法测定[17]。采用土壤试剂盒(南京建成生物工程研究所)测定土壤酶活性。
1.5 DNA提取、16S基因扩增及Illumina Miseq测序
土壤DNA采用土壤DNA试剂盒[生工生物工程(上海)股份有限公司]从每个土壤样品中分离得出,使用分光光度计(A260/A280)对DNA浓度和质量进行定量,确保所有DNA产量为1.0~2.5 mg。利用细菌偏向引物338F(5′-ACTCCTACGGAGCAGCAGCAG-3′)和806R(5′-GGACTACHVGGTWTCTAAT-3′)扩增细菌16S rRNA基因V4~V5区的468 bp片段。用含5×FastPfu缓冲液4 μL、dNTPs(2.5 mmol·L-1)2 μL、正向引物(5 μmol·L-1)0.8 μL、反向引物(5 μmol·L-1)0.8 μL、FastPfu聚合酶0.4 μL、DNA模板10 ng、BSA 0.2 μL加ddH2O至20 μL反应混合物进行PCR扩增。PCR参数:95 ℃持续3 min(变性);95 ℃持续30 s(变性)27个循环;55 ℃持续30 s(退火);72 ℃持续45 s(延长);72 ℃持续10 min和10 ℃直到停止(ABI GeneAmp®9700)。产物经琼脂糖凝胶电泳纯化后回收。PCR产物在上海美吉生物Illumina Miseq平台测序(http://www-MaigrBio.com)。97%的同源性下进行操作分类单元(OTU)分析,物种分类信息利用Silva(Release128 http://www.arb-silva.de)数据库比对,数据处理图形生成均在Majorbio I-Sanger云平台(www.i-sanger.com)进行。
1.6 统计分析
采用Excel 2010进行数据计算;采用SPSS 15统计分析,P<0.05有统计学意义。采用单因素方差分析(ANOVA)和Fisher最小显著差异(LSD)检验进行多重比较。
2 结果与分析
2.1 轮茬作物对苜蓿产量的影响
前3次刈割,苜蓿产量轮茬玉米(T1)>轮茬高粱(T2)>苜蓿连作(CK),且差异显著(P<0.05);第4次刈割,轮茬玉米(T1)与轮茬高粱(T2)差异不显著(P>0.05),连作苜蓿(CK)显著小于轮茬处理(P<0.05);第5次刈割,轮茬玉米(T1)产量显著高于苜蓿连作(CK)及轮茬高粱(T2)(P<0.05),连作苜蓿(CK)与轮茬高粱(T2)差异显著(P<0.05);最终,苜蓿年产量轮茬玉米(T1)>轮茬高粱(T2)>苜蓿连作(CK),且差异显著(P<0.05),轮茬玉米(T1)及轮茬高粱(T2)后苜蓿产量分别是苜蓿连种(CK)的1.27和1.13倍(表1)。
2.2 轮茬作物对土壤化学性质变化的影响
与苜蓿连作(CK)相比,轮茬高粱(T2)能显著提高土壤pH及土壤OM(P<0.05),但轮茬玉米(T1)处理土壤pH及OM并没有显著变化(P>0.05);与连种(CK)相比,轮茬玉米(T1)土壤总氮含量增加到了2.16 g·kg-1,轮茬高粱(T2)有所增加,但并不显著(P>0.05);轮茬玉米(T1)土壤中有效磷含量变化不显著(P>0.05),轮茬高粱(T2)显著降低(P<0.05);轮茬玉米(T1)速效钾含量从连作苜蓿(CK)的173.60 mg·kg-1提高到489.17 mg·kg-1,差异显著(P<0.05),轮茬高粱(T2)提高到322.03 mg·kg-1,差异显著(P<0.05)(表2)。
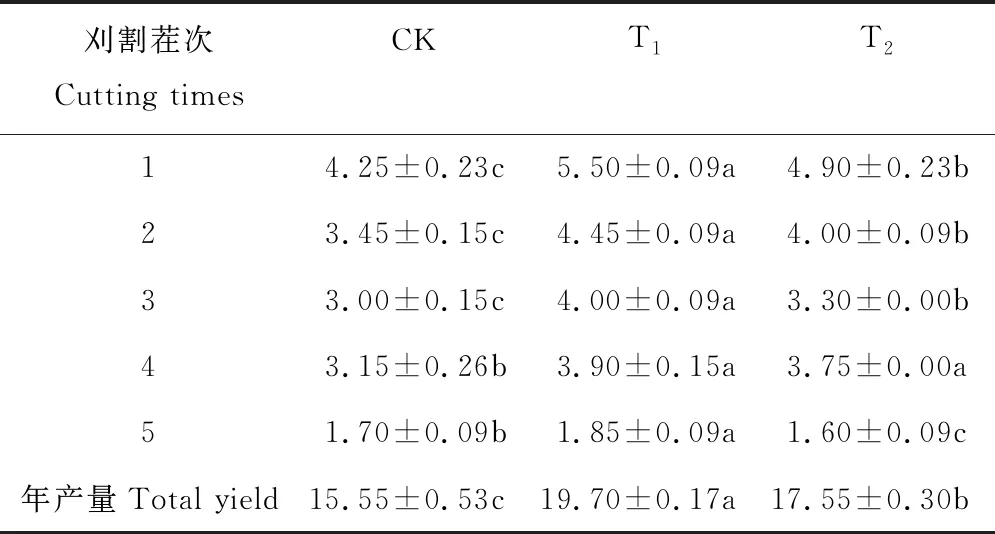
表1 轮茬作物对苜蓿产量及产量构成的影响Table 1 Annual yield and cutting yields in each treatment (t·hm-2, dry matter) (mean±SE, n=3)
注:CK,对照;T1,轮茬玉米;T2,轮茬高粱。同行不同小写字母表示差异显著(P<0.05)。
Note: CK, Control; T1, Incorporation of maize into the rotation; T2, Incorporation of sorghum into the rotation. Different lowercase letters within the same row indicate significant differences at the 0.05 level.

表2 轮茬作物对土壤化学性质变化的影响Table 2 Chemical characteristics of soil in each treatment in 2015 (mean±SE, n=3)
注: CK,对照;T1,轮茬玉米;T2,轮茬高粱。同列不同小写字母表示差异显著(P<0.05),下同。
Note: CK, Control; T1, Incorporation of maize into the rotation; T2, Incorporation of sorghum into the rotation.Different lowercase letters within the same column indicate significant differences at the 0.05 level, the same below.
2.3 轮茬作物对土壤容重变化的影响
与苜蓿连种(CK)相比,轮茬玉米(T1)和轮茬高粱(T2)均在一定程度上降低了土壤容重,使得土壤更为疏松,在0~10 cm土层,轮茬高粱(T2)根部土壤容重显著小于苜蓿连种(CK)及轮茬玉米(T1)土壤,苜蓿连种(CK)及轮茬玉米(T1)差异不显著(P>0.05);在10~20 cm土壤容重苜蓿连种(CK)>轮茬玉米(T1)>轮茬高粱(T2),轮茬高粱(T2)与苜蓿连种(CK)和轮茬玉米(T1)差异显著(P<0.05),苜蓿连种(CK)与轮茬玉米(T1)差异不显著(P>0.05);在20~30 cm土层,轮茬玉米(T1)及轮茬高粱(T2)均显著小于苜蓿连种(CK)(P<0.05),可以看出轮茬高粱(T2)在降低土壤容重方面表现更为突出,在一定程度上缓解因连种引起的土壤板结(表3)。
2.4 轮茬作物对土壤酶活性的影响
从土壤酶活性(表4)可以看出,与连种苜蓿(CK)相比较,轮茬玉米(T1)和轮茬高粱(T2)土壤碱性磷酸酶的活性差异不显著(P>0.05);轮茬玉米(T1)与轮茬高粱(T2)土壤过氧化氢酶活性的值相差不大,分别为173.40和172.03 μmol H2O2·g-1·24 h-1,但均高于苜蓿连种(CK)土壤144.11 μmol H2O2·g-1·24h-1, 且达到显著水平(P<0.05);蔗糖酶在轮茬玉米(T1)土壤中最低,轮茬高粱(T2)与连种苜蓿(CK)差异不显著(P>0.05);轮茬玉米(T1)和轮茬高粱(T2)土壤中脲酶活性分别提高达到了2385.07及2287.28 μg NH3-N·g-1·24 h-1,均显著高于连作苜蓿(CK)(P<0.05)。
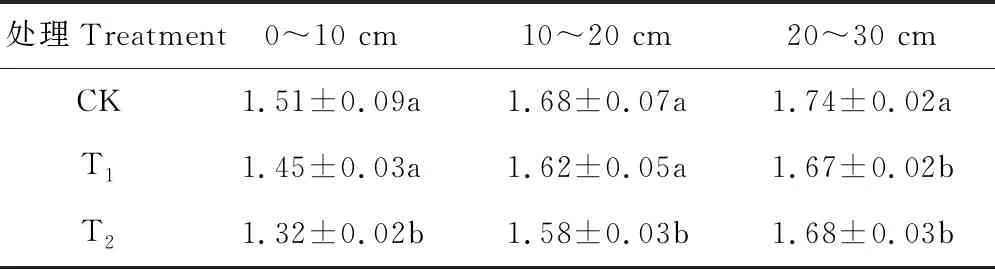
表3 轮茬作物对土壤容重的影响Table 3 Bulk density of soil in each treatment (g·cm-3) (mean±SE, n=3)

表4 轮茬作物对苜蓿土壤酶活性的影响Table 4 Enzyme activities in each treatment (mean±SE, n=3)
2.5 轮茬作物对土壤细菌群落多样性的影响
2.5.1不同轮茬作物对根际土壤细菌群落结构特征的影响 利用Illumina Miseq分析,检测细菌群落结构在不同轮茬处理下的变化,在不同处理下,变形菌(Proteobacteria)、拟杆菌(Bacteroidetes)、厚壁菌(Firmicutes)均为前3位相对丰度高于10%的优势细菌门。从连作苜蓿(CK)土壤细菌组成看,变形菌、拟杆菌、厚壁菌所占比值分别为36.19%、21.06%、13.35%(图1a),在轮茬玉米(T1)中所占比值分别为32.10%、19.49%、12.16%(图1b);在轮茬高粱(T2)中所占比值分别为27.86%、18.40%、11.02%(图1c)。在3个处理中,丰度最高的前3位的细菌,变形菌相对丰度CK(36.19%)>T1(32.10%)>T2(27.86%);拟杆菌相对丰度CK(21.06%)>T1(19.49%)>T2(18.40%);厚壁菌 CK(13.35%)>T1(12.16%)>T2(11.02%)。占比最高前3位细菌相对丰度均在苜蓿连作(CK)土壤高于轮茬玉米(T1)高于轮茬高粱(T2),从而可以看出,土壤中细菌群落结构轮茬高粱(T2)土壤最为稳定,轮茬玉米(T1)次之,苜蓿连作(CK)土壤最不稳定。
2.5.2不同轮茬作物对根际土壤细菌群落多样性的影响 9个土壤样品中提取DNA,获得了16S基因的PCR产物,PCR产物的测序结果显示总共获得4675个细菌物种OTU(图2)。连作苜蓿(CK)、轮茬玉米(T1)、轮茬高粱(T2)的16S rDNA个数分别为2847、3947、3545,分别代表连作苜蓿(CK)、轮茬玉米(T1)及轮茬高粱(T2)土壤中细菌物种OTUs;连作苜蓿(CK)专属物种OTU 198个,轮茬玉米(T1)专属物种OTU 310个,轮茬高粱(T2)专属物种OTU 677个;基于平均丰度的轮茬玉米(T1)和轮茬高粱(T2)共有物种OTU 3015个,连作苜蓿(CK)和轮茬高粱(T2)之间的共有物种OTU 2429个,连作苜蓿(CK)和轮茬玉米(T1)的共有物种OTU 2394个,连作苜蓿(CK)、轮茬高粱(T2)和轮茬玉米(T1)共有物种OTU 2174个,轮茬高粱(T2)>轮茬玉米(T1)>苜蓿连作(CK);从而看出,轮茬玉米(T1)和轮茬高粱(T2)处理根际土壤微生物活性显著高于连作苜蓿(CK),物种OTU数量是连作苜蓿 (CK)的1.25和1.39倍。
2.5.3不同轮茬作物对根际土壤细菌群落多样性指数特征的影响 轮茬玉米(T1),轮茬高粱(T2)土壤OTU水平的ACE指数比苜蓿连作(CK)稍有提高(图3),但3者之间的差异不显著(P>0.05); Shannon指数显示,连作苜蓿(CK)和轮茬玉米(T1)差异不显著(P>0.05),而与轮茬高粱(T2)差异达到了极显著水平(P<0.001);Sobs指数显示,连作苜蓿(CK)和轮茬玉米(T1)差异不显著(P>0.05),而与轮茬高粱(T2)差异达到了极显著水平(P<0.001);Chao指数显示,连作苜蓿(CK)和轮茬玉米(T1)差异不显著(P>0.05),而与轮茬高粱(T2)差异达到了极显著水平(P<0.01)。可见,轮茬高粱(T2)显著提高了根际土壤细菌群落的多样性。
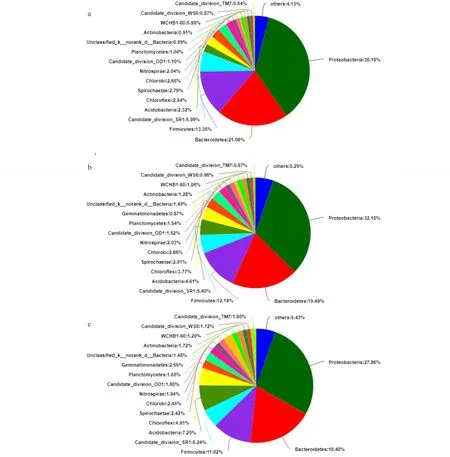
图1 不同处理土壤样品的细菌结构组成Fig.1 Bacteria community of soil samples for different treatments a: CK; b: T1; c: T2; Proteobacteria: 变形菌门; Bacteroidetes: 拟杆菌门; Firmicutes: 厚壁菌门; Candidate_division_SR1: 未分离培养细菌; Acidobacteria: 酸杆菌门; Chloroflexi: 绿湾菌门; Spirochaetae: 螺旋菌门; Chlorobi: 绿菌门; Nitrospirae: 硝化螺旋菌门; Candidate_division_OD1: 未分离培养细菌; Planctomycetes: 浮霉菌门; Gemmatimonadetes: 芽单胞菌门; Unclassified_k_norank_d_Bacteria: 未分类细菌; Actinobacteria: 放线菌门; WCHB1-60: 未分离培养细菌; Candidate_division_WS6: 未分离培养细菌WS6; Candidate_division_TM7: 未分离培养细菌TM7; Others: 所有相对丰度低的稀有细菌门的总和The sum of all rare bacterial phyla with low relative abundances.
2.5.4轮茬作物对根际土壤细菌群落丰度的影响 在细菌门水平,比较了丰度前15的不同细菌在不同处理下的丰度差别(图4)。结果显示,相较于连作苜蓿(CK),轮茬玉米(T1)和轮茬高粱(T2)土壤的变形菌均显著减小(P<0.01);而厚壁菌也是在苜蓿连作(CK)土壤中丰度最高,在轮茬玉米(T1)土壤中有所减少,在轮茬高粱(T2)土壤中丰度最低,并且3者之间变化达到显著水平(P<0.05);绿湾菌在苜蓿连作(CK)土壤种相对丰度最低,在轮茬玉米(T1)土壤中相对丰度有所增加,在轮茬高粱(T2)土壤中,相对丰度最大,3者达到了极显著水平(P<0.001);浮霉菌也是在苜蓿连作土壤中丰度最低,轮茬玉米(T1)土壤中有所增加,而轮茬高粱(T2)土壤中相对丰度达到最高值,3者达到显著水平(P<0.05);在丰度最高的前15个细菌门,其他细菌的相对丰度变化在不同处理下,差异不显著(P>0.05)。
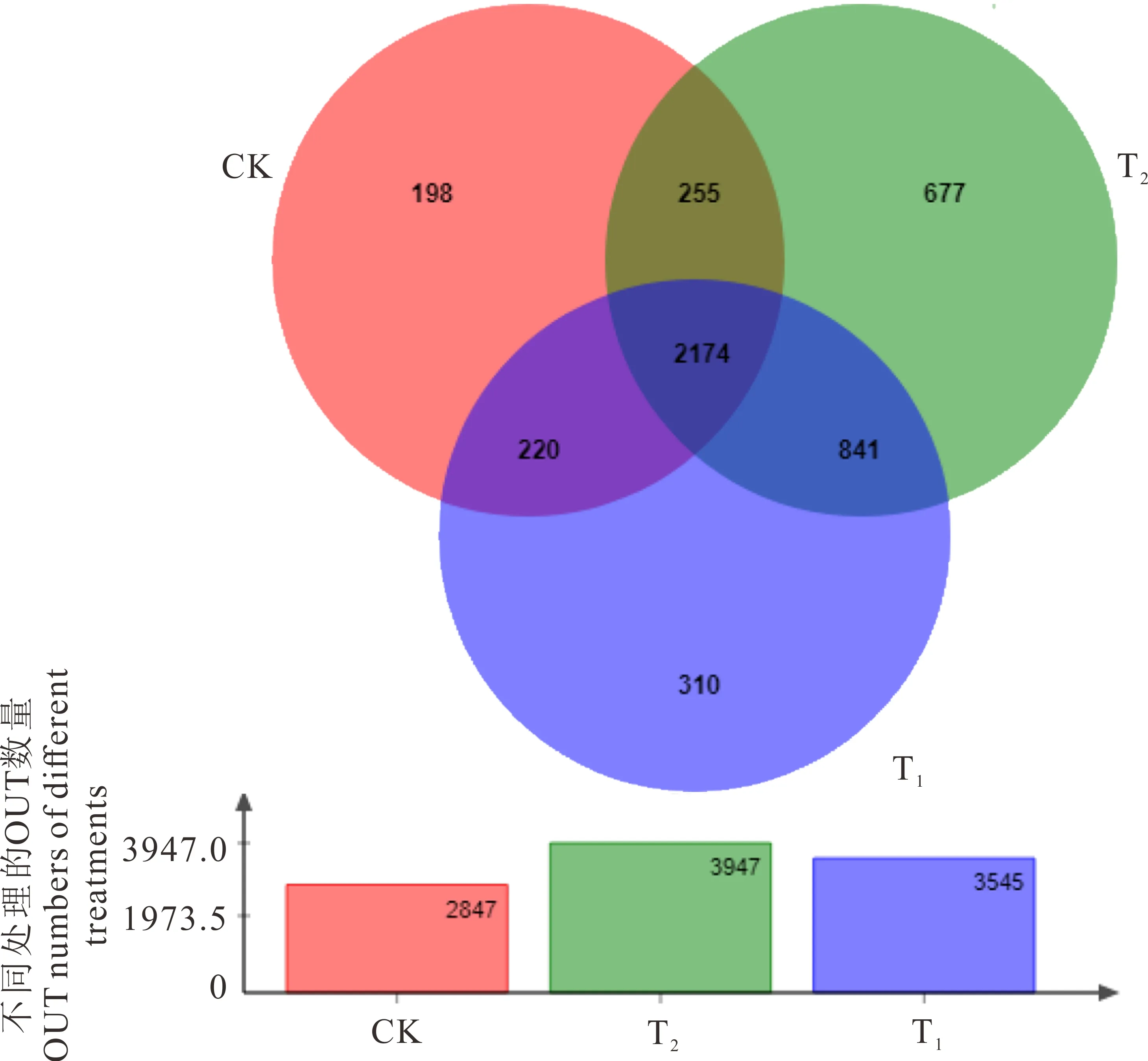
图2 不同处理在OTU水平对土壤细菌组成的影响Fig.2 Soil bacteria community composition of different treatments on OTU level
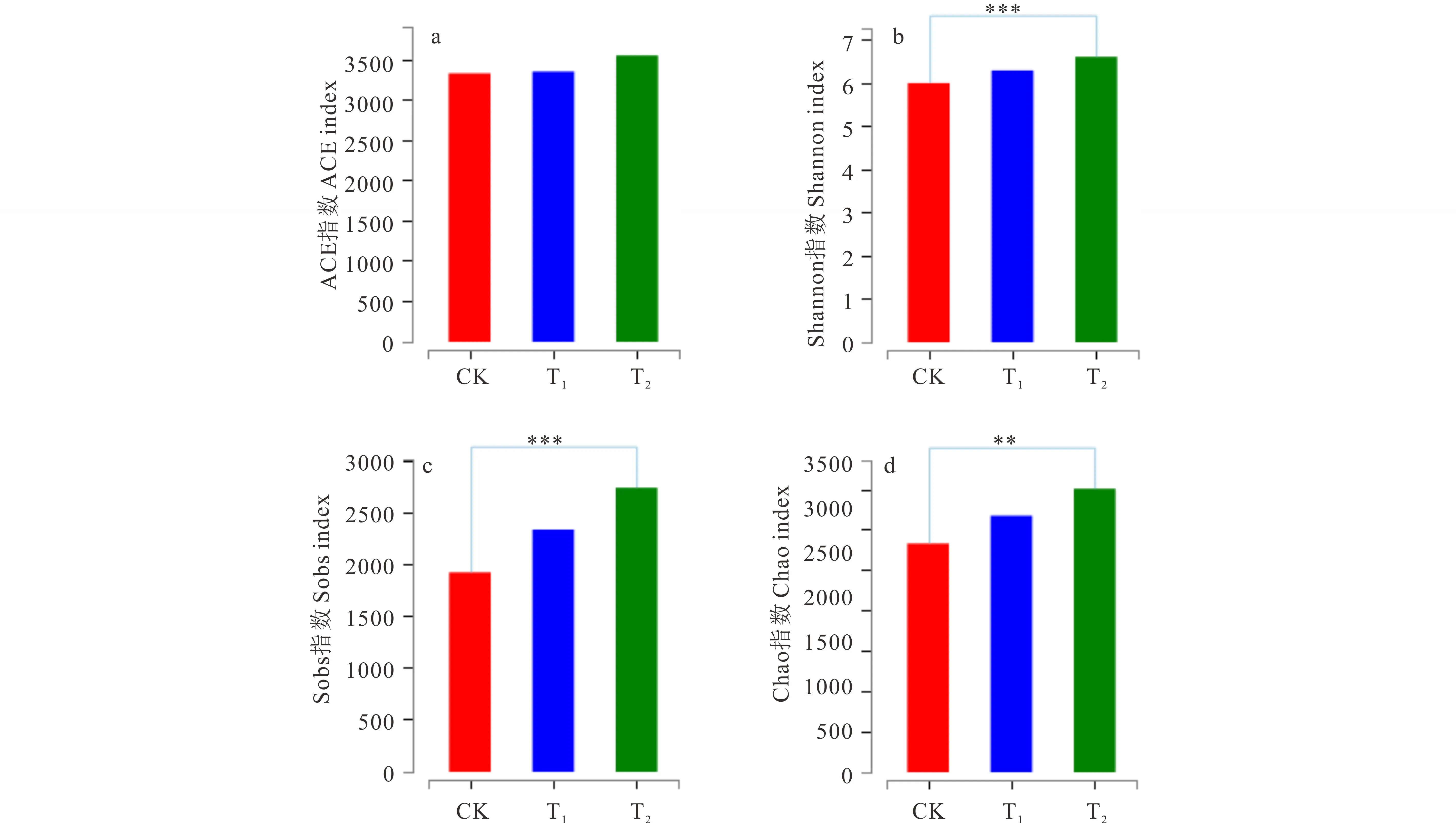
图3 不同处理在OTU水平对土壤多样性的影响Fig.3 Soil bacteria composition diversity of different treatments on OTU level *: P<0.05; **: P<0.01; ***: P<0.001. 采用t检验评估OTU水平各样本的细菌多样性,下同。The bacterial diversity of each sample on OTU level are estimated by t test, the same below.
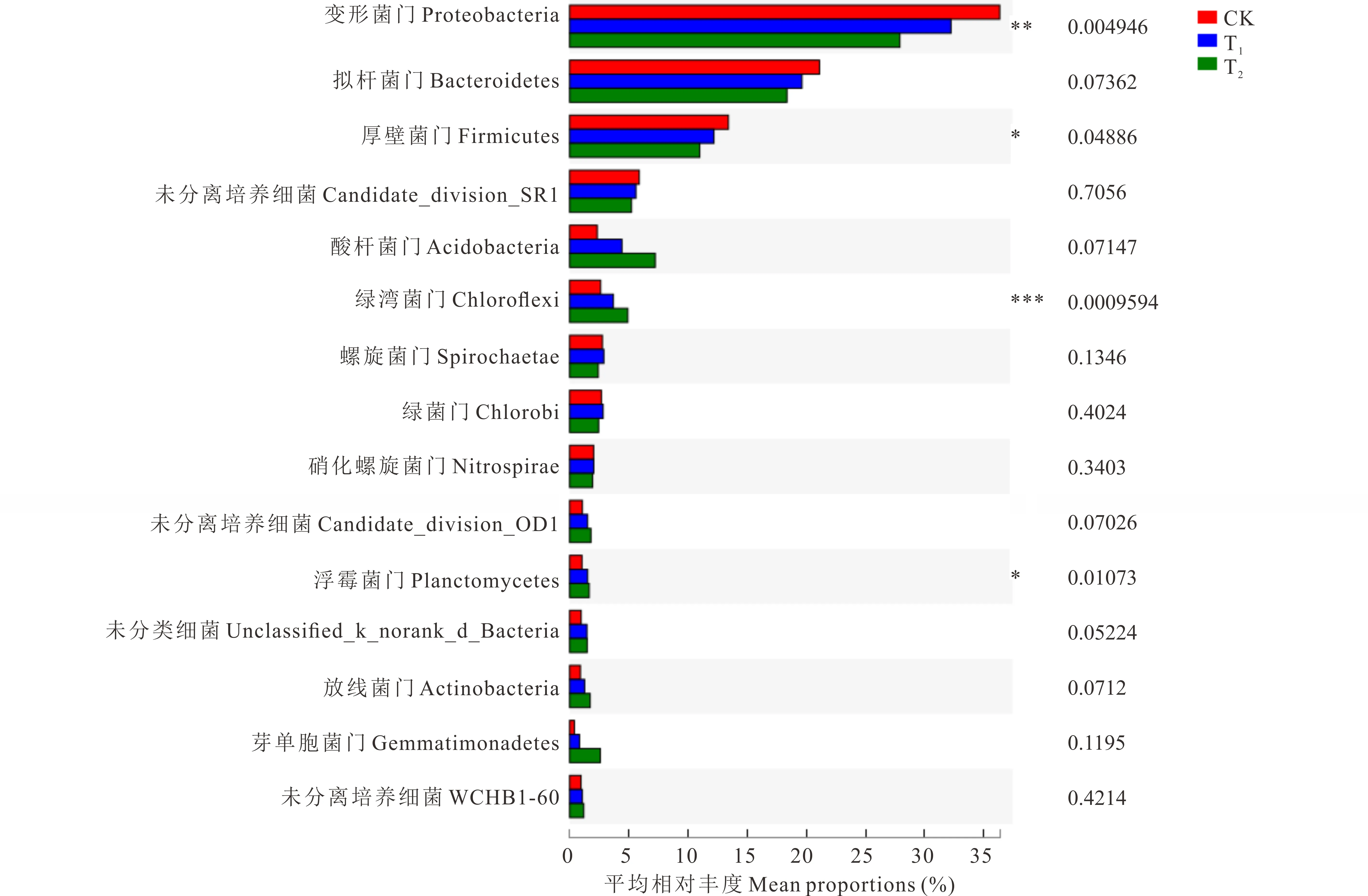
图4 不同处理细菌相对丰度(前15个门)变化Fig.4 Different bacteria abundance based on percentages on phylum level, top 15 phyla
3 讨论
3.1 不同轮茬处理对土壤理化性质变化的影响
很多作物都会表现出连作障碍,尤其豆科牧草,对农业生产造成很大的影响。轮作是一种缓解连作障碍的有效措施,能改善土壤健康状况,促进作物生长,增加产量[12,16,18-19]。以往的研究表明,土壤pH随着连作时间的延长而降低,土壤修复能力逐渐降低[20-21],pH与土壤质量呈负相关,与土壤健康呈负相关,与连作和病害呈正相关[24]。而合理的轮作能够提高连作土壤的pH,土壤pH和养分含量可通过调节土壤的生物学特性[22]来调控长期种植作物的产量。与之相似,本研究结果显示,轮茬使土壤pH有所增加。由于不同作物随养分的需求消耗并不相同,因而导致土壤残留养分差异。研究表明,与轮作相比,连作茄子(Solanummelongena)土壤有效磷、钙、镁含量显著降低[16]。土壤有机质也是影响连作土壤质量和提高作物抗病性的关键因素。Davis等[25]发现土壤有机质含量与较大的微生物生物量和较高的作物产量有关。与苜蓿连作模式相比,苜蓿不同作物轮作处理能够调整不同形式碳的含量变化[23]。本研究发现,轮茬使土壤有机质增加,使得土壤更加肥沃,总氮、速效钾含量均高于连作土壤,而有效磷含量轮茬高粱低于连作及轮茬玉米土壤,说明轮茬高粱后要适当增加施磷量。土壤容重能够反映土壤的通气性、透水性和植物根系生长的阻力状况,其数值的大小决定于土壤质地、结构和松紧程度,是土壤物理性质的一个重要指标,土壤容重越小,孔隙度越大,说明土壤发育良好,有利于植物生长[26]。本研究发现,轮茬在不同根层降低了土壤容重,疏松土壤,更有利于苜蓿的生长。
3.2 不同轮茬处理对土壤酶活性大小变化的影响
土壤酶活性是土壤生物化学过程的重要指标[10],它的大小可以反映土壤养分转化的能力,脲酶和碱性磷酸酶参与氮磷循环和有机质分解[27],过氧化氢酶是一种可以减轻生物体内毒素的氧化还原酶,蔗糖酶能够催化蔗糖水解为葡萄糖和果糖,进而提高土壤生物学活性,增加土壤营养[27]。在棉花、小麦玉米轮作中也发现轮作能够提高土壤过氧化氢酶、蔗糖酶及相关酶活性[28]。10年生苜蓿翻耕后轮作不同作物,结果显示轮作土壤酶活性均要高于连作,而且玉米、高粱轮作的土壤酶活性要高于作物连作及谷子(Setariaitalica)、马铃薯的轮作方式[18]。与轮作相比,连作茄子土壤多酚氧化酶、脲酶、碱性蛋白酶活性降低,并且不同的轮作方式,土壤酶活性的大小表现不同[16]。本研究表明,土壤脲酶活性在轮茬土壤中显著高于连作土壤,脲酶可以水解有机质分子中肽键,说明土壤中具有较强的转氮能力。轮茬高粱及玉米均显著提高了土壤过氧化氢酶活性,说明轮茬土壤减轻生物体内毒素的氧化还原酶,土壤健康状况更好。综上,轮茬处理提高了土壤酶活性,增强了土壤的氧化还原能力,提高了养分的吸收及循环水平。
3.3 不同轮茬处理对土壤细菌群落多样性变化的影响
细菌是构成土壤微生物最重要的组成部分,微生物群落变化可导致土壤代谢能力和土壤质量的变化,是土壤健康的重要决定因素,它们通过影响代谢酶在土壤中的沉积进而影响土壤的质量和功能[29],土壤细菌群落受到农业管理技术如作物连作和轮作[9,30-31]的影响[19]。同时,土壤微生物多样性在可持续农业中发挥着重要作用,因为微生物可以影响很多生化过程包括植物养分循环、土壤结构维护和农业化学降解,从而促进农业生产。通常认为:ACE(物种丰富度ACE指数)、Sobs(丰富度实际观测值)、Chao(物种丰富度Chao1指数)指数反映了群落丰富度(community richness)、Shannon指数(Shannon多样性指数)反映了群落多样性(community diversity)。群落丰富度(community richness)越高,则样本土壤微生物优势种群越突出,群落多样性指数(Shannon)越低,而多样性指数越高,优势物种丰富度越低,土壤微生物环境越稳定。研究已经表明,连作能够引起根际土壤中微生物群落的重大变化,如果土壤的微生物种群结构失衡,有害微生物会对植物生长产生负面影响,形成连作障碍[32],通过合理轮作,能够协调作物与土壤的关系,从而实现作物高产及农田生态环境的改善,维持农业的可持续生产。已有报道表明,轮作可以提高大棚黄瓜土壤的Shannon指数,轮作土壤中微生物组成及多样性均要高于连作土壤[19];10年生苜蓿翻耕后轮作不同作物,结果显示轮作土壤微生物数量显著高于连作土壤[18];小麦轮作也能够显著提高土壤微生物多样性[7];棉花-牧草-冬小麦轮作与棉花连作相比,土壤的微生物量在0~5 cm、5~10 cm、10~20 cm土层均高于连作土壤,而且轮作明显降低了土壤真菌细菌比值,提高了土壤质量[10];与轮作相比,连作茄子土壤细菌分类单元数量减少,真菌增多,多样性指数降低,轮作水稻及四季豆(Phaseolusvulgaris)、长瓜(Cucurbitaceae)较连作更有利于提高土壤肥力和土地生产力[16]。本研究采用Illumina Miseq测序法,揭示了连作和轮茬土壤中细菌群落结构组成,结果表明,与苜蓿连作(CK)相比,轮茬玉米(T1)和轮茬高粱(T2),OTU物种数量明显增加,土壤细菌种类更为丰富稳定,轮茬高粱(T2)显著提高了苜蓿根际土壤细菌群落的Shannon、Chao及Sobs多样性指数,这说明轮茬高粱后,土壤细菌群落的数量、代谢活性及多样性均升高,土壤活力增强,肥力更高,营养元素的循环代谢更为高效,而轮茬玉米虽有提高但差异并不显著,可能是由于不同轮茬作物根系分泌物的不同影响了土壤细菌群落多样性[33],而连作导致根系分泌物单一,土壤中微生物的代谢结构单一,微生物功能多样性降低,轮茬能够优化根际土壤环境,有利于多种微生物的生长代谢。本研究表明,轮茬高粱的土壤中细菌丰度及多样性要高于轮茬玉米,这与虎德钰等[18]的研究结果一致,可能是由于高粱吸收肥料水分能力较强,土壤的代谢要高于其他作物,因而土壤微生物数量要高于其他作物。本研究通过对不同细菌丰度分析表明连作及不同作物轮茬对土壤根际不同细菌丰度也有不同的影响,所以2种轮茬作物调控根际土壤微生物群落的机理各有不同。本研究发现,变形菌在细菌结构组成中,属于丰度最高的细菌,一般土壤中变形菌为土壤细菌丰度最高的细菌门类[14],连作土壤中相对丰度显著高于轮茬土壤,这可能是因为苜蓿根瘤菌属于α-变形菌,因而在苜蓿连作土壤中相对丰度最高,连作有利于苜蓿土壤中根瘤菌的成长和积累。有研究表明,随着黑胡椒(Pipernigrum)连种年限的增加,连作时间的延长,厚壁菌相对丰度逐年降低,而厚壁菌对土壤病原真菌具有拮抗作用[34],而本研究中轮茬处理土壤中,厚壁菌相对丰度低于连作土壤,造成这个不同的原因可能是作物种类的不同,苜蓿连作可能有利于厚壁菌的积累及生长。而变形菌及厚壁菌处于表达丰度最高前3位,连作土壤中高丰度表达优势物种也印证了其成为土壤细菌菌群结构的不稳定因素。而本研究显示轮茬处理土壤中绿湾菌相对丰度显著高于(P<0.01)连作土壤,而绿湾菌具有生态恢复的能力,可以增强土壤中营养物质的吸收[35],可见,轮茬土壤比苜蓿连作,优化了根际土壤环境。浮霉菌和土壤中的硝态氮的正相关性,与土壤氮代谢有密切关系[36],而本研究显示,轮茬土壤中浮霉菌相对丰度显著高于连作土壤,证明,轮茬土壤更有利于营养元素的代谢吸收。综上,轮茬土壤中细菌群落多样性的增加,使土壤菌群更为稳定健康,促生菌相对丰度增加,以及对有害病菌的拮抗作用增强,对苜蓿生长环境起到了促进作用。
4 结论
江淮地区丰富的水热资源在利于苜蓿生长的同时也造成更为严重的连作障碍,不利于农业的产业化及可持续化发展。从以上试验可以看出,轮茬提高了部分土壤酶活性,土壤细菌多样性显著增强,部分功能微生物丰度增加,进而使土壤活力增强,微生物代谢增强,功能多样性增强,促进了苜蓿产量。连作3年苜蓿-玉米-苜蓿的轮茬模式能更显著的提高苜蓿产量,而连作3年苜蓿-高粱-苜蓿的轮茬模式更有利于土壤质量的提高,因此,在江淮地区苜蓿产业化种植区域,合理轮茬能有效提高牧草产量及土壤质量,维持农田生产力,提高经济效益。
猜你喜欢
快乐作文(1.2年级)(2023年9期)2023-05-12 11:31:34
古今农业(2022年2期)2022-08-15 01:39:52
青年文学家(2022年1期)2022-03-11 12:27:39
今日农业(2021年11期)2021-11-27 10:47:17
现代畜牧科技(2021年9期)2021-10-13 06:38:52
现代畜牧科技(2021年4期)2021-07-21 06:13:04
中国化肥信息(2018年12期)2018-03-01 03:12:54
新农业(2017年24期)2018-01-30 08:37:26
中国三峡(2017年4期)2017-06-06 10:44:22
为了孩子(3~7岁)(2016年6期)2016-05-14 03:09:17
