
马铃薯油菜素内酯信号激酶基因StBSKs的克隆与序列分析
2019-08-29 07:26陈春艳
浙江农业学报 2019年8期
马 杰,郑 好,周 平,陈春艳,吴 瑞,马 维,宋 雷,孙 勃,*
(1.四川农业大学 园艺学院,四川 成都 611130; 2.毕节市农业科学研究所,贵州 毕节 551700)
马铃薯(Solanumtuberosum)是茄科茄属一年生草本植物[1],原产于南美洲安第斯山区,自17世纪传入我国后,就已成为我国传统的、主要的粮食作物之一,也是重要的蔬菜、饲料和工业原料,以及多用途的高产、高适应性农作物[2]。马铃薯具有效益高、耐瘠薄、产业链长、易于加工、营养价值高等优势[3]。油菜素内酯(brassinolide,BR)是广泛存在于植物中的相似于动物甾醇类激素的一种天然产物[4]。大量植物生理与分子生物学研究结果表明,油菜素内酯在植物的正常生长发育过程中是不可缺少的[5],许多植物生理学家已将其列为植物的第6大类激素。BR普遍存在于植物中,在植物的不同器官,如根、茎、叶、花粉、雌蕊、果实和种子中均有分布[6]。BR参与调控植物生长发育、植物与环境间的相互作用,并能增加作物产量、改善品质和增强作物抗逆性[7],还能够提高作物的抗冷性[8]、抗热性[9]、抗旱性[10]等。BR之所以具有这些功能,可能是因为BR可以激活植物中的氧化酶保护系统,从而尽快消除植物体内由于逆境而产生的过多有害自由基,提高植物抵抗逆境的能力[11],以此增强植物抗性。拟南芥中存在12种BSK蛋白激酶,这12种都属于受体细胞质蛋白激酶第12家族(RLCK-XII)。BSK1是BR信号通路中的一个重要的信号转导激酶,它与定位在膜上的BRI1结合并在BR诱导下,作为BRI1的底物被激活,从而将BR信号传导到下游,介导BR诱导的抗氧化防护酶活性的增强[4]。
目前,关于BR信号转导途径的研究大都仅限于拟南芥信号转导途径组分的同源基因克隆和相应突变体的分析[12],BSK基因在拟南芥[13]、水稻[14]、玉米[4]等中有部分研究,但在马铃薯中的研究尚未见报道。本试验克隆了马铃薯StBSK基因家族的7个成员,为进一步研究马铃薯StBSK基因的调控功能奠定了分子基础,也为明确StBSK基因在增加马铃薯植株的胁迫耐受能力中的作用提供了理论依据。
1 材料与方法
1.1 试验材料
本试验以马铃薯栽培品种宣薯2号为材料,2017年9月将其原原种块茎播种于花盆中,定期进行水肥管理,21 d后剪取马铃薯植株幼苗的3~5片真叶,液氮速冻后于-80 ℃超低温冰箱中保存,用于总RNA提取。
1.2 试验试剂
反转录试剂盒、DL 2 000 DNA marker购于大连宝生物公司,TransStart FastPfu Fly DNA Polymerase聚合酶、Trans1-T1克隆感受态细胞、克隆载体pEASY-Blunt等购于北京全式金生物技术有限公司,氨苄青霉素(Amp)、SanPrep柱式DNA胶回收试剂盒等购于生工生物工程(上海)股份有限公司。
1.3 总RNA提取与反转录cDNA合成
采用改良的CTAB法[15]提取马铃薯的RNA,电泳检测RNA的完整性,并利用核酸蛋白仪测定RNA的浓度和纯度(D260/D280=1.8~2.0为宜)。选取质量较好的RNA依据TaKaRa反转录试剂盒说明书合成cDNA模板,所得产物于-20 ℃保存。
1.4 基因克隆
根据NCBI已公开的拟南芥等BSK基因序列,设计马铃薯BSKs基因特异性引物(表1),引物由生工生物工程(上海)股份有限公司合成。PCR反应体系(40 μL):pfuFly聚合酶0.8 μL,上下游引物(10 μmol·L-1)各1.6 μL,cDNA模板1.6 μL,Buffer 8 μL,dNTPs(2.5 mmol·L-1)3.2 μL,ddH2O 23.2 μL。其中,StBSK1、StBSK4、StBSK6、StBSK7的反应条件为:94 ℃ 2 min;94 ℃ 20 s,53 ℃ 20 s,72 ℃ 25 s,40个循环;72 ℃ 10 min。StBSK2、StBSK3、StBSK5的反应条件为:94 ℃ 2 min;94 ℃ 20 s,55 ℃ 20 s,72 ℃ 25 s,40个循环;72 ℃,10 min。
扩增产物经1.0%琼脂糖凝胶电泳检测合格后,使用SanPrep柱式DNA胶回收试剂盒纯化回收产物。将产物连接至平末端pEASY-Blunt克隆载体上,转入大肠埃希菌Trans1-T1感受态细胞中,涂布于含IPTG、Amp和X-gal的LB平板上,倒置于37 ℃生化培养箱培养8~12 h,待长出明显而又未相互重叠的单菌落时,将平板放置于4 ℃数小时至显色完全。挑取10个生长状态良好的白色单菌落于1 mL LB培养基中扩大培养。以菌液为模板进行PCR扩增,将电泳检测条带长度为1 500 bp左右的菌液送生工生物工程(上海)股份有限公司测序。
表1 马铃薯StBSKs基因克隆引物
Table 1 The cloning primer ofStBSKsinSolanumtuberosum

引物名称Primer name引物序列Primer sequence (5’-3’)StBSK1-FGGGTTATCATGGGTTGTTGTCAATCTTCStBSK1-RAGGGTATTATCAAGATGCACGTCCAStBSK2-FAGCTGATGAAAAATGGGCTGTTTACAGStBSK2-RCCAGAATCAGTTACGCCAACTGTTTAACStBSK3-FGGGGTAATAGGATGGGCTGTGAGStBSK3-RTCGCCACTTTCAGGAATTTGTGTTCStBSK4-FGTTGGATAGTGTGATGGGCTGTGAAAStBSK4-RGCCACTTTCAAGAAGATGTGTTTTTCTCStBSK5-FTGCTCTGAAATGGGTGGTCGTTCStBSK5-RTTAATTTTTGCTCCTTTTGCCTTCCAACTStBSK6-FTGTTTTCATTTTCACTGTCAAATGGGTGCStBSK6-RAATGCAAATCTATACAGTTTCAGTTTTTGTStBSK7-FATGGGTGGTCGTTGTTCCAATTTATGStBSK7-RTTATTGTCTCATTCTTCTGAAGTCCAAGG
1.5 生物信息学分析
利用ExPASy(https://web.expasy.org/protparam/)在线软件预测StBSKs基因的相对分子质量、等电点、不稳定指数等理化性质;由TMHMM(http://www.cbs.dtu.dk/services/TMHMM/)软件分析和推测蛋白质跨膜结构;通过WoLF PSORT(https://wolfpsort.hgc.jp/)软件进行亚细胞定位预测;使用NCBI-BLAST程序(https://blast.ncbi.nlm.nih.gov/Blast.cgi)结合DNAMAN 6.0软件进行同源序列比对和保守结构域分析;用MEGA 6.0软件进行系统进化树的构建[16-17]。
2 结果与分析
2.1 马铃薯StBSKs基因的克隆
经电泳检测可明显地观察到28S和18S条带,且28S条带的亮度在18S条带亮度的2倍以上(图1),D260/D280比值为1.9,表明马铃薯总RNA质量合格,可用于后续克隆试验。将StBSKs基因的PCR扩增产物经1%琼脂糖凝胶电泳检测后,分别得到7个长度约为1 500 bp的特异性条带(图2),这些条带大小与预测片段一致。阳性克隆测序得到7条长度分别为1 497、1 479、1 464、1 461、1 476、1 476和1 476 bp的序列,并将其分别命名为StBSK1、StBSK2、StBSK3、StBSK4、StBSK5、StBSK6和StBSK7。
2.2 生物信息学分析
2.2.1 StBSKs蛋白理化性质分析和亚细胞定位预测
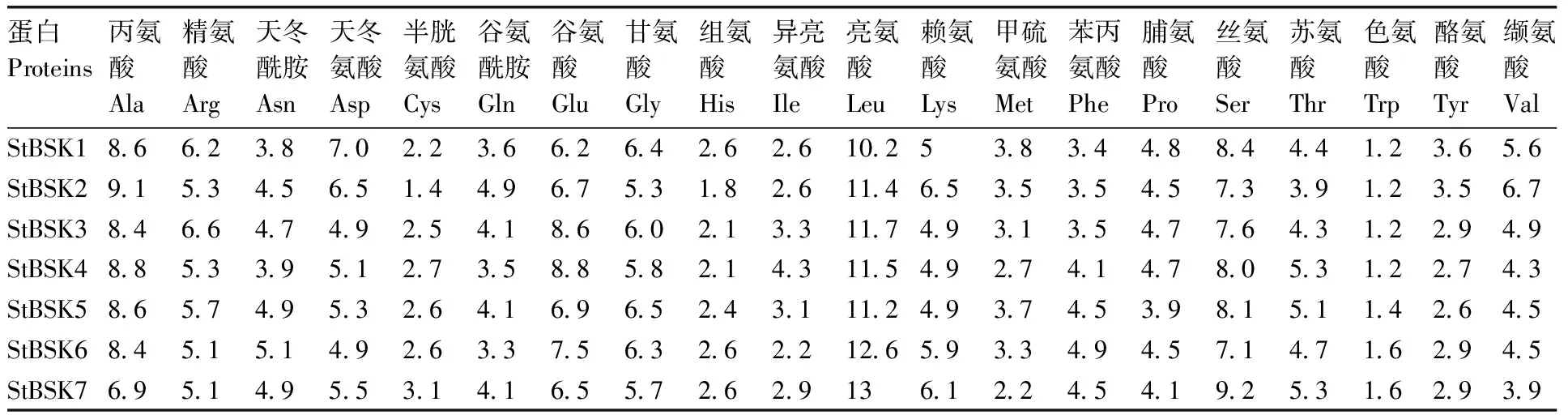
马铃薯StBSKs基因所编码的氨基酸序列的理化分析结果如表2所示。不稳定指数小于40为稳定蛋白,大于40为不稳定蛋白,由此可知,StBSK1、StBSK3、StBSK4和StBSK5为不稳定蛋白,而StBSK2、StBSK6和StBSK7为稳定蛋白。平均疏水指数均为负值,表明其均为亲水性蛋白。StBSKs蛋白的氨基酸组成具体见表3;SignalP、TMHMM等软件分析发现,StBSKs蛋白均不存在信号肽结构和跨膜结构域,均为非分泌蛋白或膜外蛋白。亚细胞定位表明,StBSK1主要位于细胞核,StBSK2、StBSK3、StBSK4主要位于细胞质,StBSK5、StBSK7主要位于叶绿体,StBSK6主要位于线粒体。
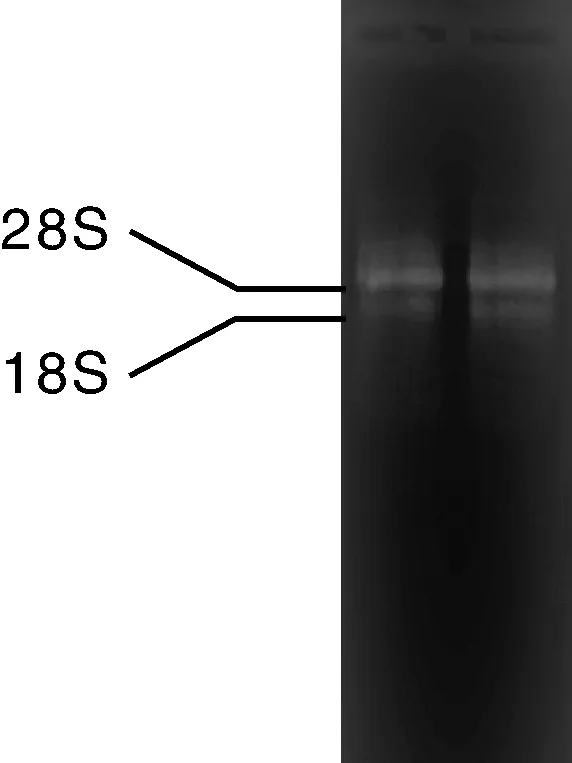
图1 马铃薯总RNAFig.1 Total RNA extracted from Solanum tuberosum

M, DL 2 000 marker; 1, StBSK1; 2, StBSK2; 3, StBSK3; 4, StBSK4; 5, StBSK5; 6, StBSK6; 7, StBSK7。图2 马铃薯StBSKs基因PCR扩增产物Fig.2 PCR amplification products of StBSKs in Solanum tuberosum
表2 马铃薯StBSKs基因家族生物信息学分析结果
Table 2 Bioinformatics analysis ofStBSKsgene family inSolanumtuberosum

基因Genes长度Length/bp氨基酸数Aminoacidnumber理论分子量Calculatedmolecularmass/ku理论等电点Isoelectricpoint(pI)总原子个数Totalnumberof atoms不稳定指数Instabilityindex脂肪指数Liposolubleindex平均疏水指数Averagehydrophobicindex亚细胞定位Subcellularlocalization转运肽长度Transitpeptidelength分子式MolecularformulaStBSK1149749855648.005.77771541.7075.06-0.421细胞核Nucleus32C2435H3815N685O750S30StBSK2147949255242.975.83773439.2583.29-0.378细胞质Cytoplasm4C2441H3856N672O741S24StBSK3146448754732.225.64763146.5781.17-0.407细胞质Cytoplasm21C2399H3794N676O735S27StBSK4146148654237.635.14756444.7483.17-0.288细胞质Cytoplasm4C2391H3760N650O737S26StBSK5147649154930.435.90762544.1777.15-0.336叶绿体Chloroplast39C2410H3776N674O734S31StBSK6147649155245.086.05769636.9179.33-0.326线粒体Mitochondria4C2452H3818N670O727S29StBSK7147649155306.886.37770737.2080.10-0.381叶绿体Chloroplast25C2444H3824N674O739S26
表3 马铃薯StBSKs氨基酸组成比例
Table 3 Proportions of amino acid composition of StBSKs inSolanumtuberosum
%
2.2.2 马铃薯StBSKs蛋白结构域与氨基酸序列
保守结构域和序列比对结果表明,StBSKs蛋白为RLCK-Ⅻ类超家族,均含有典型的N端激酶结构域PKc(putative kinase catalytic)、C端三羧氨酸重复TPR(tetratricopeptide repeats)结构域,StBSKs成员间N端存在较大差异。7个StBSKs蛋白成员间的氨基酸序列相似性达到75.89%,成员间两两互相比对,相似性在55.49%~87.27%,其中,StBSK3和StBSK4氨基酸序列的相似性最高(87.27%)(图3)。
2.2.3 马铃薯StBSKs蛋白系统进化树
StBSKs的氨基酸序列与辣椒(Capsicumannuum)、芝麻(Sesamumindicum)、中粒咖啡(Coffeacanephora)、黄灯笼辣椒(Capsicumchinense)、浆果状辣椒(Capsicumbaccatum)、木犀榄(Oleaeuropaeavar.Sylvestris)、番茄(Solanumlycopersicum)等多种植物的BSK蛋白氨基酸序列一致性高达90%以上,与中华猕猴桃(Actinidiachinensisvar.Chinensis)、木薯(Manihotesculenta)、毛果杨(Populustrichocarpa)等植物的BSK氨基酸一致性达80%以上。13种不同植物与马铃薯StBSKs蛋白的系统进化树分析结果表明,StBSKs大致可分为2大分支,马铃薯StBSK1、StBSK2被聚在第一大分支,其中,StBSK1、StBSK2分别与同为茄科的辣椒、野生烟草(Nicotianaattenuata)聚在2个小分支,这一分支还包括了同为大戟科的木薯(Manihotesculenta)和橡胶树(Heveabrasiliensis);马铃薯StBSK3和StBSK4与同为茄科的黄灯笼辣椒聚在一个小分支,StBSK5和StBSK6与同为茄科的浆果状辣椒聚在一个小分支,StBSK7与同为茄科的番茄聚在一个小分支,他们共同聚在第二大分支。表明,同科物种间的BSK亲缘关系更近,其中,StBSK3和StBSK4、StBSK5和StBSK6关系最近(图4)。
3 结论与讨论
油菜素内酯是一种类似动物甾醇类激素的植物激素,广泛存在于各种植物的不同器官,具有调控植物生长发育、增强作物抗逆性的生物学功能。在BR的信号通路中,BSK是一个重要的信号转导激酶,将BR信号传导到下游。本试验首次分离了马铃薯StBSK基因家族的7个成员。氨基酸序列比对和系统进化树结果表明,7个马铃薯StBSK成员均与茄科的物种聚为同支,说明BSK在进化过程中与同科物种间的亲缘关系较近。StBSKs基因含有典型的保守结构域PKc、三羧氨酸重复结构域,PKc位点在氨基酸序列73~307,三羧氨酸重复结构域位点在390~489,这2个保守域分别在细胞分裂、增殖、凋亡、分化过程中发挥作用,还能介导蛋白与蛋白间的互作[18]。7个马铃薯StBSKs氨基酸序列存在一定差异性,且N端保守性较差。StBSKs氨基酸序列富含丝氨酸和苏氨酸,含少量的酸性氨基酸,这与转运肽的性质相符。亚细胞定位分析表明,这些马铃薯StBSKs蛋白可能主要位于细胞质、细胞核、叶绿体和线粒体,因此推测这些StBSKs蛋白的转运肽类型可能不同。

图3 马铃薯StBSKs氨基酸序列比对Fig.3 Alignment of deduced amino acid sequences of StBSKs in Solanum tuberosum
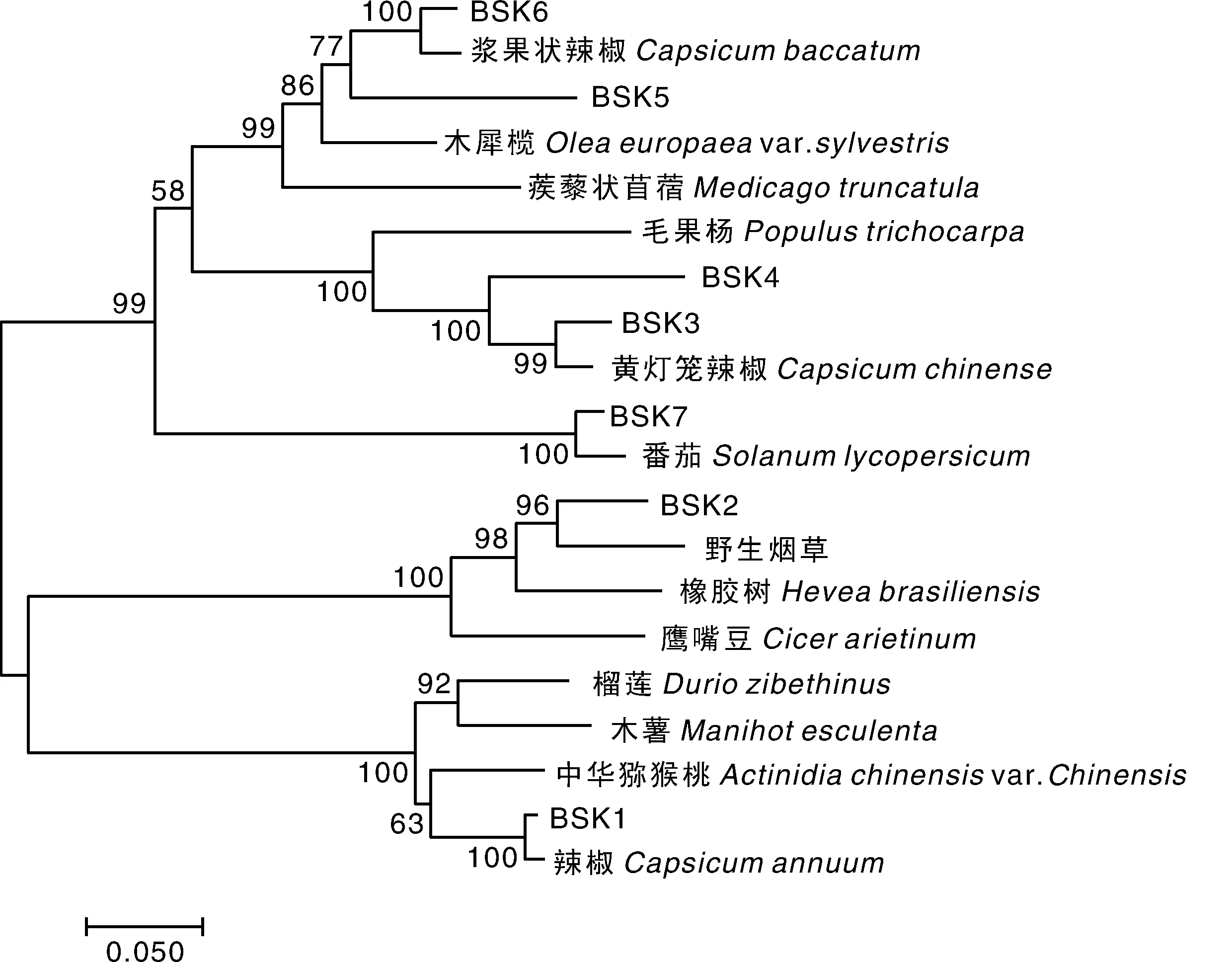
图4 马铃薯和其他植物的StBSKs蛋白系统进化树Fig.4 Phylogenetic tree analysis of StBSKs in Solanum tuberosum and other plants
BSK基因家族成员在不同植物中的数目和功能存在异同。拟南芥中有12个BSK基因,玉米中有14个BSK基因,玉米中ZmBSK1蛋白可能在质膜系统中起感应与传导信号的作用;ZmBSK5可能与AtBSK3、AtBSK4、AtBSK6、AtBSK7、AtBSK8功能相似,能够引起植物表型变化;ZmBSK9可能与AtBSK1作用相同,均能结合受体BRI1,调节玉米的BR信号转导途径等;ZmBSK2.1与AtBSK1同源,其可能参与免疫反应(PAMP-triggered Immunity, PTI)的调节[19]。综上,推测StBSKs基因在马铃薯生长发育过程与抗逆境胁迫中发挥重要调控作用,还参与BR信号的感应和传导作用。
猜你喜欢
环球时报(2022-09-20)2022-09-20
湖北农业科学(2022年11期)2022-07-18
河北农业大学学报(2022年2期)2022-04-26
食品安全导刊(2021年21期)2021-08-30
湖南饲料(2021年3期)2021-07-28
兽医导刊(2020年13期)2020-12-31
今日农业(2020年24期)2020-12-15
实用肿瘤学杂志(2020年4期)2020-12-08
小资CHIC!ELEGANCE(2015年14期)2015-09-23
小资CHIC!ELEGANCE(2015年15期)2015-09-01
