
羊肚菌菌丝间融合与不融合现象研究
2019-08-20 03:02贺新生王亚洲谭仁豪竹文坤
食药用菌 2019年4期
贺新生 赵 苗 王亚洲 张 能 谭仁豪 王 银 竹文坤 陈 波
羊肚菌菌丝间融合与不融合现象研究
贺新生 赵 苗 王亚洲 张 能 谭仁豪 王 银 竹文坤 陈 波
(西南科技大学生命科学与工程学院,四川 绵阳 621010)
用平板、空试管、断培养基等方法培养羊肚菌菌丝,在显微镜下原位直接观察菌丝间的融合和非融合现象。观察六妹羊肚菌、梯棱羊肚菌、七妹羊肚菌、Mel-21和秋天羊肚菌、Mes-16、Mes-19、Mes-20、Mes-21等9个物种,共100多个菌株菌丝间2 000多个接触点;并历经6年时间,用300多个菌株在国内外100多个地点超过500公顷面积进行出菇试验。结果发现:同一单孢菌株的菌丝分枝间有少量部位的可自体融合,融合点不产生新的分枝菌丝;同一物种的不同单孢菌株间菌丝不融合,同一物种的多孢菌株培养物内部相互不融合;不同物种间的菌丝都不融合;不融合的菌丝接触点出现II型、X型、Y型、T型、π型等状况交叉而过,细胞接触而不融合;未见菌丝间的有性融合;菌丝接触后出现死亡、缠绕、卷曲、打结、膨大等现象;羊肚菌菌丝体纯培养物可以形成分生孢子和菌核。出菇试验结果表明,腐生型羊肚菌物种的出菇率,超过200个单孢菌株100%,超过100个多孢菌株100%,超过50个纯组织分离物小于50%,超过30个组孢混合菌株小于50%;出菇菌株在形态和产量有显著差异。共生型羊肚菌的菌种都不出菇。能出菇的同一物种的多孢菌株出菇后表现出子实体形态多样性,不同物种混合播种后出菇的子实体为原来物种的形态特征。结论:羊肚菌单孢培养物菌丝间不发生有性融合,单个孢子自身可孕,完全能够单孢出菇。
羊肚菌菌丝;单孢出菇;有性融合;显微镜原位直接观察;菌丝空试管培养法
菌丝融合是所有丝状真菌的物种在其生长发育过程中普遍发生的一种现象。菌丝融合的类型分为:无性融合、有性融合。无性融合是同一细胞发育的菌丝相邻分枝之间发生的自体融合;同一物种的不同菌株的菌丝之间会发生无性融合;菌丝在无限的无性繁殖过程中相邻菌丝之间产生分枝或直接接触,相互融合使得菌丝之间形成三维立体的网络,有利于占据更大的生存空间和营养源,有利于对营养的吸收与运输、信息传递和菌丝体、子实体的生长。有性融合是在两个不同性别的单孢菌株菌丝之间,菌丝接触后相互融合,形成具有全部基因组的新细胞,新细胞萌发出新菌丝,完成有性生殖,成功繁殖下一代,形成大量有性孢子,如子囊孢子、担孢子、接合孢子。无性融合、有性融合都可用于物种的鉴定,有性融合多用于杂交育种。
在异宗结合真菌的有性生殖过程中,2个不同性别的可亲和的单孢单核菌丝相互接触后,细胞发生有性融合,形成一个新的双核异核细胞,由此细胞开始萌发出新的可以完成有性生活史的双核异核菌丝,原来的单核菌丝逐渐死亡或不再生长。显微镜下可以直接观察到融合以后的细胞上长出新的菌丝的现象。异宗结合的担子菌中单个单核菌丝不会出菇,2个或多个同性别的单孢培养物混合培养也不会出菇,只有2个或2个以上的不同性别的培养物混合培养后才会出菇。因此对一个物种是否是异宗结合,是否可以进行杂交育种的判断法则是:单孢培养物不出菇,可亲和的菌丝间细胞发生融合,融合以后的新菌丝具有锁状联合,可形成新的子实体和有性孢子。
羊肚菌属于子囊菌门,子囊菌的菌丝可能有不结实和可结实的菌丝,因为缺乏担子菌门菌丝的锁状联合和其他形态学特征,纯培养物难形成子实体,无法根据菌丝形态来判断子囊菌的培养物的可结实性和不同性别的菌丝之间是否交配成功。要证明单孢菌株菌丝之间有交配行为,必须要有显微镜下原位直接观察的证据,确证菌丝之间有普遍的有性融合现象,否则无法判断结论是否正确。用压片法观察菌丝,制片过程中容易打乱菌丝的自然排列状况;用扫描电子显微镜观察菌丝也不能打乱菌丝的原始排列状况,在技术上很难做到。
羊肚菌的菌丝很粗,一般直径有5~20mm,有3~5或更多级的分枝,分枝级数越多尖端菌丝越细;老菌丝有一定的颜色,隔膜和分枝非常容易识别,都会形成特殊的菌核和分生孢子。由于菌丝直径差异明显,羊肚菌菌丝很容易与其他的丝状真菌区分开,如各种霉菌、担子菌等,在低倍光学显微镜下可直观地识别。要观察菌丝之间的融合和非融合现象,可采用空试管培养、培养皿平板培养孢子萌发或菌丝,可在显微镜下原位直接观察、拍照试管壁或打开的培养皿正面、反面,很容易得出正确的结论。也可以用载玻片直接培养法、盖玻片插片法培养,取下载玻片、盖玻片在显微镜下原位观察。
羊肚菌的菌丝在培养基中的空间分布情况为:在培养基内部的基内菌丝,表面的匍匐菌丝,培养基上空的气生菌丝;菌核有培养基表面的菌核,气生菌丝的菌核;分生孢子分布在气生菌丝体尖端。要观察菌丝之间是否融合就要分别观察基内菌丝、匍匐菌丝、气生菌丝之间是否有接触和融合、非融合的现象发生。
羊肚菌的每一个子囊中有8个子囊孢子,颜色、大小、形态差异不显著,单个孢子、各种菌丝培养物都是多核体,在形态上没有显著性的差异,无法判断其遗传性别。其子囊孢子极易萌发,在有水的条件下萌发率一般超过90%,存活时间很长,一般不会很快死亡,新生的菌丝在形态上没有差异。要判断单个孢子的结实性,可以通过单孢分离进行出菇试验来加以证明。
羊肚菌的菌种很容易获得,分离的方法很多,如纯组织分离、单孢分离、多孢分离、组孢混合分离、土壤或基物菌丝分离等。各种分离物的培养特征没有多大的差异,大多数能够形成菌核,培养时间超过20天后在显微镜下可以观察到分生孢子的形成。组织分离有膨大细胞分离,紧密组织分离,膨大细胞和紧密组织混合分离等方法,分离部位有菌盖、盖柄结合部位、菌柄组织等。组孢混合分离是取子实体表面的产孢组织,从成熟新鲜子实体上取组织,就有菌盖的菌肉组织和子囊孢子,萌发出的培养物是组孢混合分离物;取成熟的干子实体表面组织,孢子已经成熟,孢子萌发比菌肉组织快24~48小时,应是多孢分离物;取幼嫩子实体表面的组织,没有成熟的孢子,是组织分离物。要确定是否出菇、出菇量的高低、质量的好坏等必须进行出菇试验。
为确定羊肚菌单孢菌株之间菌丝是否能够发生菌丝的有性融合、可否进行杂交,我们特设计了几种显微镜下原位直接观察的方法,对大量的培养物作了多年的研究,并进行出菇试验,对结果进行证实。
1 材料与方法
1.1 供试菌株
试验用菌株是从9个物种的新鲜子实体或风干子实体上分离所得,包括单孢分离物、单孢+单孢组合培养物、多孢分离物、纯组织分离物、组孢分离物等。试验的9个物种均进行过分子系统学鉴定确认。
(1)可出菇的物种。六妹羊肚菌:学名M. Kuo, in Kuo, Dewsbury, O'Donnell, Carter, Rehner, Moore, Moncalvo, Canfield, Stephenson, Methven & Volk,104(5): 1170 (2012)。M5原始标本来自四川省广元市青川县青溪镇,200个分离物。M909原始标本来自四川省阿坝州九寨沟县,80个分离物。M1018原始标本为四川栽培菌株,100个分离物。G19原始标本为四川栽培菌株,30个分离物。
梯棱羊肚菌:学名M. Kuo, O'Donnell & T.J. Volk, in Kuo, Dewsbury, O'Donnell, Carter, Rehner, Moore, Moncalvo, Canfield, Stephenson, Methven & Volk,104(5): 1172 (2012)。Mma原始标本来自新疆的马鞍菌子实体,组织分离物。Mjt原始标本来自四川栽培菌株,40个分离物。Mb原始标本来自巴基斯坦野生标本,20个分离物。
七妹羊肚菌:学名M. Kuo, in Kuo, Dewsbury, O'Donnell, Carter, Rehner, Moore, Moncalvo, Canfield, Stephenson, Methven & Volk,104(5): 1171 (2012)。M7原始标本来自北川县栽培菌株。30个分离物。
Mel-21:四川省绵阳市安州区桑枣镇野生标本,5个分离物。
(2)不出菇的菌种。秋天羊肚菌:学名Masaphy & Clowez, in Clowez,. 126(3-4): 238 (2012) [2010]。野生标本来自西南科技大学校园,30个分离物。
野生羊肚菌:Mes-16、Mes-19、Mes-20、Mes-21,野生标本来自金堂县竹篙镇、中江永安镇、南部、平武锁江镇、平武平通镇、平武坝子乡、广元朝天区等地,各10个以上的孢子分离物。
1.2 培养基
水琼脂培养基、PDA培养基、PDA加富培养基[1~7]。
1.3 菌种分离方法
[1~3]的方法,得到试验用菌株。
1.4 菌丝观察方法
培养皿培养直接观察法:孢子接种在有培养基的培养皿中;培养物带培养基的菌丝块接种在无培养基的培养皿中,接种块间距1~2 cm,培养温度22~25 ℃,菌丝萌发接触后进行观察。把培养皿打开皿盖,培养基朝上,放在光学显微镜载物台上,先用4倍物镜对焦,再用10倍、20倍、40倍、100倍物镜观察和拍照。移动培养皿或试管,进行多点观察。培养基朝下,直接观察培养皿背面。
空试管培养直接观察法:将试管塞上橡皮塞后灭菌,冷却后在试管中部接入2块带培养基的菌丝培养物,间距1~2 cm,25 °C培养,菌丝接触把试管放在显微镜载物台上,显微镜4倍物镜调焦观察,再用10倍物镜观察。
断培养基观察法:将培养皿中央的培养基挖去1~2 cm的长条或圆条,在两侧接上不同的菌株。或将试管中的斜面培养基挖断并移动,间隔1~2 cm,分别在2块培养基上接种不同菌株。培养2~4天后,两侧的菌丝在断面上接触,用显微镜直接观察培养皿或试管壁。
观察层面:基内菌丝,培养基表面贴生的匍匐菌丝,培养基上空的气生菌丝。
观察点:菌丝相互接触的点、线,菌丝分枝顶端,气生菌丝尖端,超过2 000个观察点。
观察材料:单孢菌丝>200个菌株,同种或异种的单+单组合培养物>50个菌种,同种或异种的多孢菌株>50个菌株,纯组分菌株>50个菌株,组孢混合>30个菌株等。
1.5 菌丝融合与不融合的判断
在培养皿中观察菌丝相互接触处,在接触点调节显微镜焦距,多焦距立体观察。
菌丝融合:菌丝接触点细胞无隔膜,接触处不膨大或异形。沿菌丝的生长方向移动载物台,确认融合的2个分枝是否发源于同一根菌丝。变化焦距,观察融合点是否有新菌丝形成。
不融合:菌丝接触后距离几乎为0或1~2mm,有或无明显的隔膜,有或无膨大或异形细胞,有或无细小的分枝。变化焦距,观察融合点是否有新分枝菌丝形成。
1.6 出菇试验
将分离得到的各种培养物,扩大培养制作原种和栽培种,在大田进行出菇试验。方法参考文献[1~7]。时间:2013—2018年。地点:四川省、重庆市、云南省、贵州省、河南省、河北省、北京市、甘肃省、新疆维吾尔自治区、福建省、湖北省、陕西省、安徽省、湖南省等,美国、法国、澳大利亚等100多个点。累计面积超过500公顷。
2 结果与分析
2.1 菌丝融合
结果如图1、表1所示。在水、水琼脂培养基或有营养物质的培养基中,孢子在24小时内萌发,大多数孢子粉的萌发率超过90%,新鲜组织块在24~48小时萌发,干组织块在48小时以后萌发。菌丝萌发以后,很快产生多级分枝,一般有4~6级。孢子萌发的单孢菌株、纯组织分离菌株的菌丝产生分枝,相邻菌丝之间产生的分枝相互接触、或一根菌丝的分枝与另外一根菌丝接触后,有部分接触点能够融合,即自体融合或无性融合。自体融合率小于接触点总数的25%。接触点细胞壁会溶解,无隔膜出现。超过75%的接触点菌丝相互不融合。同一个菌株的不同接种块萌发的菌丝之间接触后会发生融合,即无性生长的融合。所有的融合点都没有观察到新分枝菌丝的发生。

图1 孢子萌发、多级分枝和自体融合
所有的多孢、混合培养物的同一根菌丝的分枝在少数接触点上有自体融合现象,沿菌丝生长方向进行源头追溯观察,发现都是同一根菌丝产生的不同分枝。没有观察到有性融合。
2.2 菌丝不融合
所有的单孢菌株、同或异种单孢间、同或异种多孢菌株、纯组织分离菌株、组孢混合菌株等的培养物的匍匐菌丝接触点不融合的情况复杂多样,相互接触后都会出现下列几种情况。
II型:相邻的2根或更多数量的菌丝平行排列,菌丝间隔距离为0~2mm或更远,菌丝同向或反向平行排列,长达2~5个细胞长度,即使距离为0,各自都有自己独立的细胞壁,各自尖端的菌丝又相互分开生长在不同的空间区域。3根以上的平行排列成为菌丝束。如图2左所示。
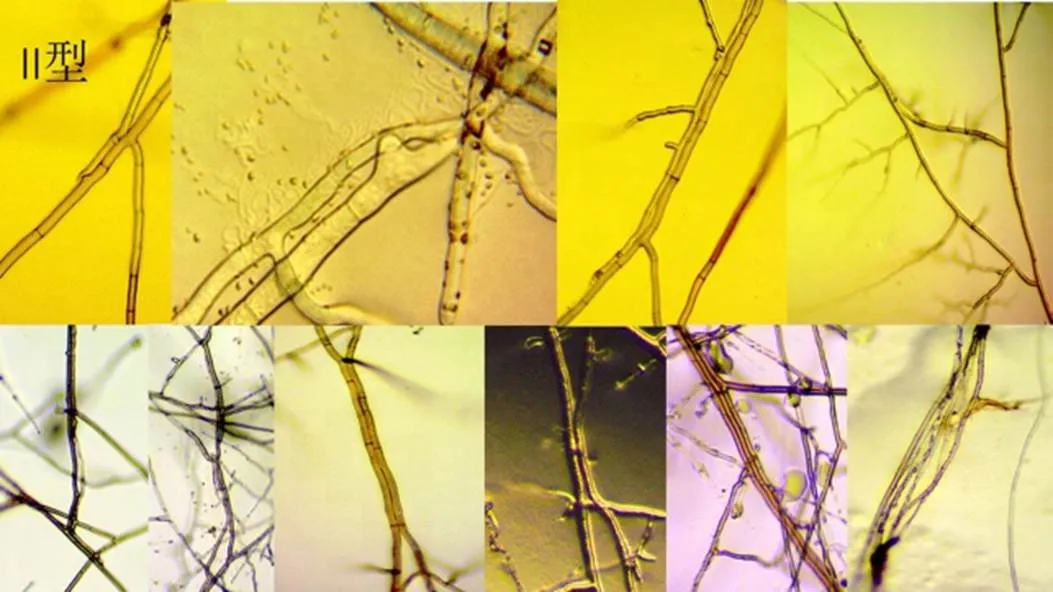
图2 菌丝不融合:II型、X型排列
X型:相邻的2根相交的菌丝呈X型、十字型交叉而过。交叉重叠处单独或各自长出短小分枝,有一根菌丝细胞在接触处明显膨大,都有自己独立的细胞壁,变化显微镜的焦距完全可以清楚观察到(图2右)。
Y型:2根相交的菌丝呈Y字形交叉而过。重叠处各自长出短小分枝,都有自己独立的细胞壁(图3左)。半Y型:2根相交的菌丝呈半Y字形交叉而过。重叠处各自长出短小分枝,都有自己独立的细胞壁。
T型:一根菌丝垂直接触于另外一根菌丝,顶端不产生新的尖端菌丝,接触处有各自的细胞壁。接触处细胞稍膨大。如图3右所示。

图3 菌丝不融合:Y型、T型交叉
π型:2根菌丝接触后,一根菌丝产生2个或2个以上的分枝绕过所接触的菌丝,菌丝细胞壁不融合(图4左)。绕过:菌丝交叉处一根菌丝与另外一根菌丝平贴一小段,再绕过去,继续生长。
细胞膨大:羊肚菌菌丝在培养基中容易产生连续或不连续的膨大细胞,膨大后比原菌丝直径增加0.5~2倍(图4右)。S型菌丝:菌丝波浪状、卷曲状,不直。在有其他杂菌存在的培养物中容易观察到。
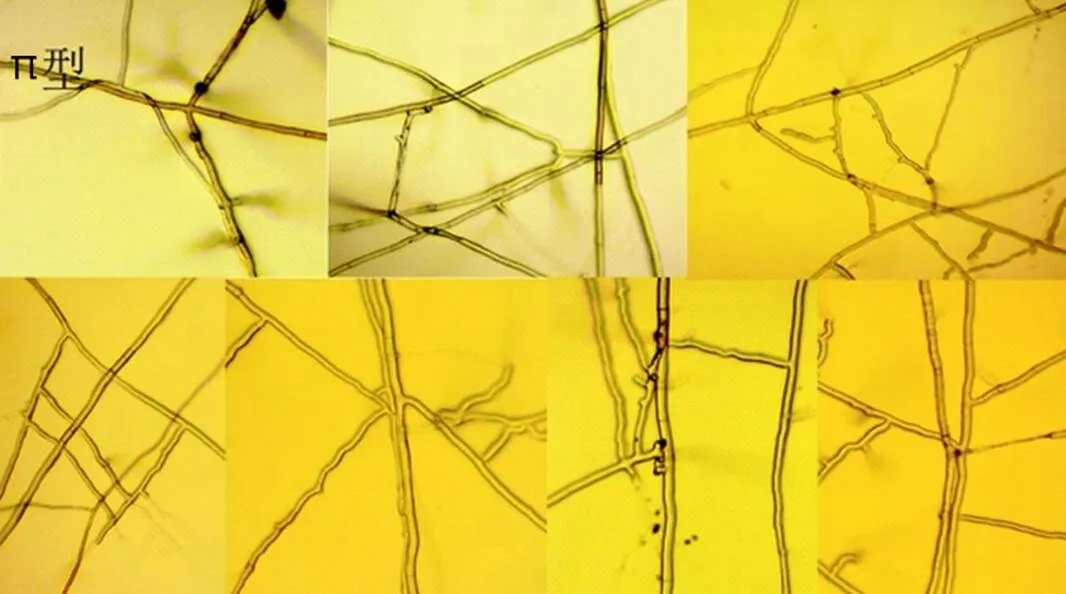
图4 菌丝π型排列和细胞膨大
2.3 菌丝死亡
在显微镜下,各种培养物都容易观察到羊肚菌菌丝细胞死亡的情况。死亡菌丝发生的部位为菌丝尖端和菌丝中段,在气生菌丝和匍匐菌丝上都容易观察到。气生菌丝尖端容易发生相互缠绕、自身卷曲、相互打结等现象,结节以后死亡的菌丝,不再发生新的菌丝。
尖端死亡:一根菌丝与另外一根菌丝接触后,尖端菌丝成为空细胞,或表面堆积大量紫红色颗粒物,细胞干瘪或变成空腔,然后死亡(图5左)。
中段死亡:一根菌丝与另外一根菌丝交叉而过以后,继续生长,接触段的2~5个细胞形成空腔细胞,死亡(图5右)。
气生菌丝相互接触后发生缠绕、卷曲和打结现象。缠绕:最细的分枝菌丝常常缠绕在较大的菌丝上,缠绕的圈数有1圈至多圈。气生菌丝最细的分枝在尖端接触后也会相互缠绕(图6左)。卷曲:菌丝顶端呈圆形卷曲1圈或多圈,呈圆盘状(图6右)。打结:细小的分枝菌丝相互缠绕后成为一个小的菌丝团(图6右)。

图5 菌丝尖端死亡和中段死亡

图6 菌丝缠绕、卷曲、打结
2.4 无性繁殖体
各种羊肚菌菌丝纯培养物中都容易观察到其无性繁殖体:分生孢子梗和分生孢子、假菌核。
分生孢子梗和分生孢子:所有的培养物中,在培养时间超过20天的老培养物上容易观察到菌丝形成的分生孢子梗和分生孢子。培养时间短的培养物中很难观察到(图7左)。

图7 羊肚菌纯培养菌丝上的分生孢子(左)和菌核(右)
假菌核或菌核:在所有的培养物中,肉眼或显微镜下可以观察到大小不等的菌丝扭结块即假菌核,稍坚硬或柔软,系菌丝在一个节点大量发生短分枝相互缠绕、或密集排列、膨大或不膨大、集群而成(图7右)。菌核常常发生在培养基表面的匍匐菌丝、气生菌丝上,贴生于培养基表面或试管壁上。基内菌丝上没有观察到菌核的发生。
2.5 出菇试验
2013—2018年在国内外多地进行出菇试验和推广应用。结果(表1)表明,单孢菌株、单+单菌株、种内多孢菌株、种间多孢菌株都能够出菇,出菇率为100%,可以多年重复出菇,小区试验产量在0.5~2.4 kg/m2范围,大面积推广应用的产量达到100~498.5 kg/667m2。各物种的产量依次为:梯棱羊肚菌>六妹羊肚菌>七妹羊肚菌>Mel-21。单孢菌株的形态、质量都比较一致,多孢分离物、混合播种的子实体形态变化大、同一栽培基地同样栽培方法的产量差异显著。
纯组织分离的菌株、组织和孢子混合分离的菌株出菇率小于50%,有的菌株可以连续多年出菇,有的产量不稳定,一年高一年低,小区试验产量在0.1~1.2 kg/m2范围。能够出菇的菌株在子实体形态和产量都有显著差异。如菌盖的圆整程度,有的很圆,有的呈明显的扁圆形;顶部有尖顶、圆顶、方顶等。能出菇的同一物种的多孢菌株出菇后子实体形态表现出多样性,不同物种混合播种后出菇的子实体呈现原物种的形态特征。
秋天羊肚菌、Mes-16、Mes-19、Mes-20、Mes-21等共生型的羊肚菌,组织分离和孢子分离菌种都可以产生菌核和分生孢子,但是都不出菇。

表1 羊肚菌菌丝间的融合与不融合及出菇试验结果
注:*为不能出菇的物种不出菇。表内“+”表示有,“﹣”表示无;“纯单”为一个孢子的纯培养物,“单+单”为2个不同的单孢培养物,“多孢”为多个孢子混合培养物,“纯组”为纯组织分离的培养物,“组+孢”为组织分离与孢子分离的混合培养物。
3 讨 论
显微镜原位直接观察羊肚菌菌丝,发现能够出菇或不能够出菇的菌种的每一个单孢菌株,部分菌丝的分枝、顶端之间接触后容易发生融合。接触后细胞壁溶解,结合处不形成隔膜。基内菌丝不容易发生融合。很多匍匐菌丝、气生菌丝相互接触后也不发生融合,而是出现菌丝相互交叉排列现象。气生菌丝会发生卷曲、缠绕、打结等现象。所有单孢培养物在通气良好的培养皿、棉塞试管培养物中都容易形成菌核。纯单孢培养物培养时间超过20天以后,在显微镜下能够观察到气生菌丝尖端形成的分生孢子梗和分生孢子,但肉眼无法看到。数百个单孢菌株的出菇试验和生产应用发现,能出菇的腐生型菌种的单孢菌株100%都可以出菇,子实体形态一致,没有完全不出菇的现象发生,菌株间、物种间的产量有明显差异。
没有观察到同一物种不同单孢菌株间菌丝发生有性融合的现象,表明同种之间单孢菌株杂交的可能性非常小;不同物种的单孢菌株之间菌丝发生融合的现象没有观察到,表明异种之间杂交的可能性也非常小。
同种、异种的多孢菌株的培养物菌丝间没有观察到不同菌丝之间的有性融合现象。在水琼脂培养基上,菌丝接触后尖端菌丝大多数变成空腔菌丝,表面堆积大量紫褐色物质。栽培菌株出菇后表现出形态学上的多样性,有圆形、椭圆形、扁平形、片状等,菌株间产量差异极显著。
纯组分菌株与单孢菌丝培养物的特征相似。能够人工栽培的物种部分菌株会出现不出菇或难出菇的情况,第二年无法重复第一年的结果。组孢混合菌株与多孢培养物的状况相似。菌丝虽有融合的现象,但95%以上的接触点都是不融合。
大量试验研究结果表明,羊肚菌出菇与不出菇菌种的各类分离物菌丝间的融合与不融合规律基本相同。在不同的单孢培养物菌丝间没有观察到有性融合,羊肚菌孢子自身可孕,完全能够单孢出菇,遗传类型不属于典型的异宗结合真菌。羊肚菌单孢杂交、多孢杂交、异种杂交的可能性极低或不会发生,这与大多数研究者的结论截然不同。
考虑到多孢菌株中菌丝之间的竞争关系,相互缠绕,尖端菌丝容易死亡等情况,栽培过程中容易出现产量、形状和质量等不稳定的问题,建议生产者多采用经过出菇试验的高产单孢菌株进行规模化生产,以避免不必要的经济损失。
目前普遍采用从孢子粉的稀释液中分离单孢的方法进行单孢分离,因而获得的单孢来自于同一个子囊的概率几乎为0。所以羊肚菌孢子分离的难点,是把同一个子囊中的8个孢子分离出来,单个分开培养,以便于对它们进行遗传差异的比较研究。
参考文献
[1] 贺新生. 羊肚菌生物学基础、菌种分离制作与高产栽培技术[M]. 北京: 科学出版社, 2017.
[2] 贺新生. 野生蕈菌生物学特性与栽培技术[M]. 北京: 中国轻工出版社, 2007.
[3] 贺新生. 羊肚菌研究进展[J]. 食用菌文摘, 1993(1): 1-9.
[4] 贺新生, 张玲, 李玉律, 等. 羊肚菌菌丝体的培养[J]. 资源开发与市场, 1994(1): 23-25.
[5] 贺新生, 陈波, 刘超洋. 中国羊肚菌栽培技术新进展[C]. 中国菌物学会第六届会员代表大会(2014年学术年会)暨贵州省食用菌产业发展高峰论坛, 2014.
[6] 贺新生. 羊肚菌人工栽培技术稳定性和规范化程度有待提高[C]. 2018第三届全国羊肚菌大会资料汇编, 2018.
[7] 贺新生, 张能, 赵苗, 等. 栽培羊肚菌的形态发育分析[J]. 食药用菌, 2016(4): 222-229.
Study on the Hyphal Fusion and Non-Fusion Phenomena between Morels
He Xinsheng Zhao Miao Wang Yazhou Zhang Neng Tan Renhao Wang Yin Zhu Wenkun Chen Bo
(School of Life Science and Engineering, Southwest University of Science and Technology, Mianyang, Sichuan, 621010, China)
The hyphae of morels was cultured in flat plate, empty test tube and broken inclined medium. The fusion and non-fusion phenomenon between mycelia were observed in situ under the microscope. Including 9 species:,,, Mel-21 and, Mes-16, Mes-19, Mes-20 and Mes-21. There are more than 2 000 contact points between mycelia of more than 100 strains. After six years, mushroom production experiments were carried out with over 300 strains in more than 100 locations and over 500 hectares around the world. The results showed that the self-fusion of a few parts between the mycelial branches of the same monospora strain, and the fusion point did not produce new mycelia; the mycelia of the same species did not fuse, and the polyspore strain cultures of the same species did not fuse with each other; the mycelia of different species did not fuse; the incompatible mycelial contact points appeared: type II, type X, type Y, type T, typeπ, etc. , the cells contacted but did not fuse; the sexual fusion between mycelia was not observed. The phenomena of death, winding, curling, knotting and swelling were observed after hyphal contact. The pure culture of morels mycelium could form asexual propagates such as conidiospores and sclerotium. The results of mushroom production test showed that the mushroom production rates of saprophyticspecies were 100% of more than 200 single-spore strains, 100% of more than 100 polyspores strains, less than 50% of more than 50 pure tissue isolates and less than 50% of more than 30 mixed strains. There were significant differences in morphology and yield of the mushroom producing strains, and none of the symbioticstrains produced mushrooms. The fruiting bodies of the same strain of mushroom-producing species showed morphological diversity after mushroom-producing. The fruiting bodies of different species after mixed sowing were the morphological characteristics of the original species. CONCLUSION: Sexual fusion does not occur between any mycelia of single-spore cultures of. Morels spores are self-fertility and can produce fruiting bodies by single-spore, and the genetic type is not the typical heterothallic fungi.
hyphae; single-spore produce fruiting bodies; sexual fusion of hyphae; microscopic in situ direct observation; method of mycelium culture in empty test tube
S646
A
2095-0934(2019)04-244-09
四川省科技厅项目(2019YFN0125)
贺新生(1965.04—),男,教授,硕士导师,主要从事食药用菌生物学和开发利用方面的研究。E-mail:hexinsheng@swust.edu.cn。
猜你喜欢
果树实用技术与信息(2022年6期)2022-11-21
今日农业(2022年16期)2022-09-22
农业工程技术(2022年1期)2022-04-19
语文周报·教研版(2021年28期)2021-08-19
江苏广播电视报·新教育(2021年34期)2021-01-03
农民科技培训(2019年7期)2019-08-16
陕西农业科学(2019年4期)2019-05-13
现代农村科技(2019年11期)2019-01-06
食用菌(2018年2期)2018-01-31
益寿宝典(2018年2期)2018-01-27
