
冰草蛋白质含量等4个重要性状的QTL定位研究
2019-07-24 11:24:26杨东升于肖夏李佳奇吴国芳牛亚青青
麦类作物学报 2019年7期
杨东升,于 卓,于肖夏,李佳奇,吴国芳,牛亚青青
(内蒙古农业大学农学院,内蒙古呼和浩特 010019)
冰草(AgropyroncristatumGaertn.)别名野麦子,是小麦族(TriticeaeDumort)密丛型多年生禾草。其根系发达,具沙套,抗旱、抗寒性强,适应性广,蕴藏着麦类作物抗逆性遗传改良的重要基因资源[1-2]。冰草春季返青早,秋季枯黄晚,生长期在200 d以上,是早春和晚秋放牧的重要牧草[3]。此外,冰草叶量较大,草质好,饲用价值高,并富含蛋白质。一般拔节期鲜草蛋白质含量约17%,开花期约14%,是禾本科牧草中蛋白质含量较高的草种。
近年来,有关冰草的形态学、生物学、细胞学、抗逆性、遗传多样性和基因工程、分子标记技术等已有广泛研究,并在生产实践中得以应用[2-6]。为创制抗逆性强、品质好、产量高的四倍体杂交冰草新种质,本课题组前期将产自内蒙古锡林郭勒草原的具优良抗性基因的二倍体野生蒙古冰草(AgropyronmongolicumKeng,2n=2x=14)与引自美国加州的具优质高产基因的栽培品种航道冰草(Agropyroncristatumcv. Fairway,2n=2x=14)进行人工授粉杂交,成功获得种间杂种F1植株,并通过秋水仙碱溶液诱导分蘖苗茎尖得到了遗传稳定的四倍体杂交冰草,其鲜草产量和蛋白质含量均显著优于二倍体亲本及其杂种F1[3-5];为构建四倍体杂交冰草的分子遗传连锁图谱,经人工套袋自交得到了加倍植株的F2代分离单株群体,以此为材料建立了高密度的遗传连锁图谱[6]。
植物重要性状的QTL定位是基因图位克隆、功能解析和分子标记辅助育种研究的基础。目前,在小麦[7]、水稻[8]、玉米[9]、马铃薯[10]、大豆[11]、油菜[12]等主要农作物的重要性状QTL定位研究方面已有较多报道,而有关牧草的QTL定位研究报道甚少,仅见于黑麦草[13]、紫花苜蓿[14]、高丹草[15]、鸭茅[16]等草种。在杂交冰草QTL定位方面,本课题组李小雷[17]、姜志艳等[18]分别对二倍体和四倍体杂交冰草的株高、分蘖数及穗型等性状进行了初步定位研究;Che等[19]以杂交冰草F1群体为材料,对其穗茎节长、穗宽等6个产量相关的性状进行了QTL定位。但有关冰草品质性状,如蛋白质、脂肪含量等性状的QTL定位尚未见报道。
本试验拟在课题组已构建出的四倍体杂交冰草高密度分子遗传连锁图谱(含763个SRAP和248个SSR标记,图谱标记间平均距离2.19 cM)基础上,对经两年一点测定的粗蛋白质和粗脂肪含量、茎叶比和单株产量共4个性状进行QTL定位分析,以期为深入开展冰草品质、产量等相关性状的基因克隆、标记辅助育种等研究奠定基础。
1 材料与方法
1.1 试验材料
材料为四倍体杂交冰草(蒙古冰草×航道冰草)F2代246个分离单株及其亲本,均种植在呼和浩特市内蒙古农业大学农场的多年生草种圃内。种植行距为40 cm,株距为25 cm。生长期内,视生长状况及时灌水、施肥,满足其对水分和养分的需求。每个单株分蘖后均形成无性株系。试验地土壤为沙壤土,肥力中等,具灌溉条件。
1.2 研究方法
1.2.1 性状的测定
于2017年6月和2018年6月,分别从四倍体杂交冰草F2代分离单株形成的246个无性系及亲本材料中,各剪取盛花期有效分蘖株,测定粗蛋白质含量、粗脂肪含量、茎叶比和单株产量4个重要性状。具体方法如下:
(1)冰草粗蛋白质和粗脂肪含量的测定:于盛花期田间随机剪取四倍体杂交冰草F2单株及亲本的地上部鲜样各30株,用羊皮纸袋分装后带回实验室;105 ℃杀青30 min;60 ℃烘干48 h,晾至室温后用小型粉碎机打成粉末状、备用。将粉碎好的样品装入检测盒,用INSTALAB○R700系列近红外品质分析仪(USA)进行测定。
(2)单株产量和茎叶比的测定:于盛花期随机剪取各供试材料50株地上部植株,称重后迅速分离茎秆与叶片,并称重,计算单株产量、茎秆与叶片的鲜重比值(茎叶比)。
1.2.2 各性状的QTL定位
在我们已构建出的四倍体杂交冰草高密度分子遗传连锁图谱[6]上,利用Map QTL 4.0[20]提供的区间作图法(Interval mapping,IM)对测定的各性状数据进行全基因组QTL扫描检测。扫描步长为1.0 cM,并将连锁系数LOD(logarithm of odds)区间视为95%的置信区间[21]。依据检测结果寻找各性状确切的QTL在不同连锁群上与其紧密连锁的标记位置和数目,并确定其遗传效应、贡献率和LOD阈值。LOD阈值确定后,将各性状检测到的QTLs用Word 2003绘图软件组合绘制成定位分布图。QTL位点命名方法为:Q+目标性状英文名缩写+QTL数。
1.3 数据统计与处理
利用Excel 2003对数据进行整理,用SPSS 22.0进行表现型及其分布分析。
2 结果与分析
2.1 冰草4个性状的表型及其分析
从表1可看出,4个被测性状在双亲间均存在显著差异,其中粗蛋白质含量(Cpc)、粗脂肪含量(Cfc)和单株产量(Fwsp)3个性状在双亲间差异极显著,且在性状表型值上,父本航道冰草均高于母本蒙古冰草(茎叶比除外)。四倍体杂交冰草F2群体的粗蛋白质和粗脂肪含量、茎叶比、单株产量均出现超亲现象,表明双亲含有对这些性状变异起调控作用的基因位点。4个性状的平均广义遗传率为70.25%,其中,茎叶比的广义遗传率为81%;单株产量的广义遗传率为59%,表明其易受环境影响。说明4个被测性状是既受环境影响又受微效多基因控制的数量性状。
由图1可知,冰草4个性状的测定值均呈正态性分布,可用于QTL定位。
2.2 被测4个性状的QTL定位
由表2、图2可知,在连锁系数LOD>2.0条件下,冰草粗蛋白质含量、粗脂肪含量、茎叶比、单株产量4个性状共检测到37个QTLs,每个连锁群上平均分布2.6个。以在连锁群LG8和LG10上分布最多,各为5个QTL;在连锁群LG5和LG6上分布最少,各为1个QTL。各性状QTL的遗传贡献率为6.1%~37.8%,其遗传效应、数目及作用方式有一定差异。
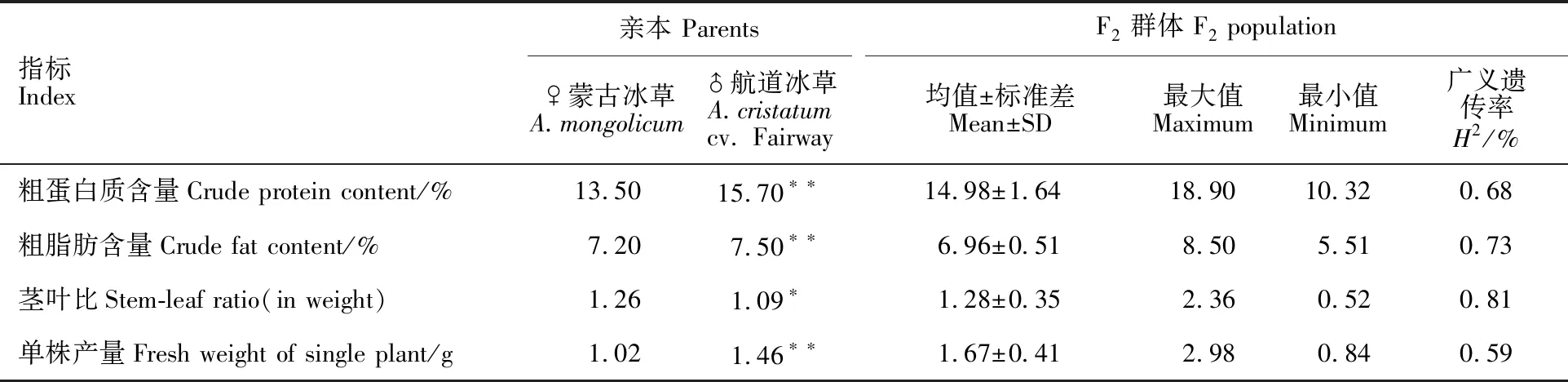
表1 冰草F2群体4个性状表现型分析Table 1 Phenotyping analysis of the four traits in F2 population of wheatgrass
*和**分别表示双亲间差异达0.05和0.01显著水平。
* and ** represent significant difference between parents at 0.05 and 0.01 levels, respectively.
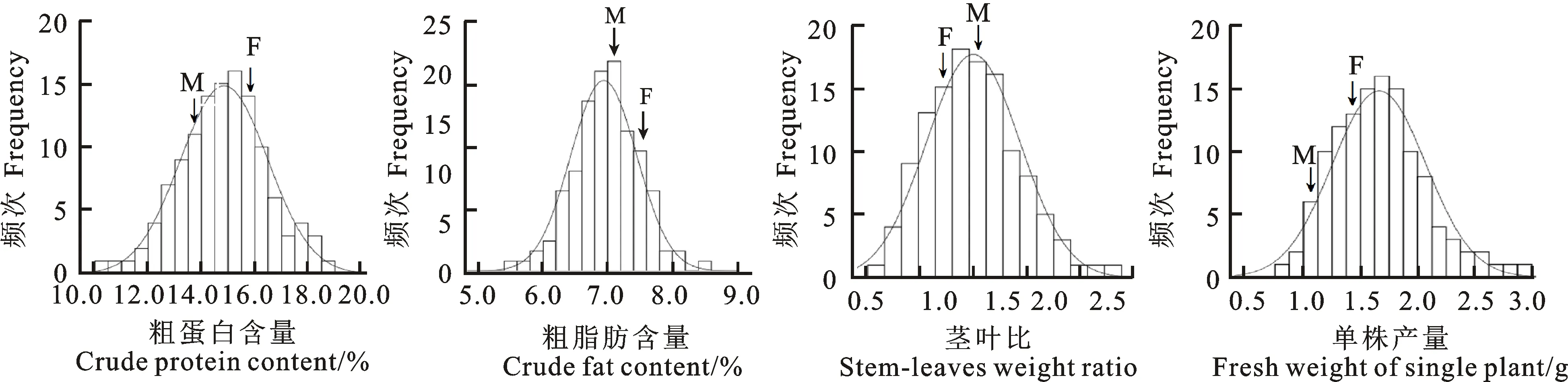
M和F分别表示母本蒙古冰草和父本航道冰草。
M and F representA.mongolicumKeng(female) andA.cristatumcv. Fairway(male),respectively.
图1 F2群体各性状的频数分布
Fig.1 Frequency distribution of each trait of F2population
2.2.1 粗蛋白质含量的QTL
检测到与冰草粗蛋白质含量相关的QTL共6个,分别是Qcpc1、Qcpc2、Qcpc3、Qcpc4、Qcpc5、Qcpc6,其分布在连锁群LG1、LG8、LG10和LG11上,遗传贡献率为6.1%~15.7%。贡献率最高的Qcpc1位于LG1的102.1 cM处,与SRAP标记m7e8-154共分离;贡献率最低的Qcpc3位于LG8的84.3 cM处,与SRAP标记m4coe9-58共分离。
2.2.2 粗脂肪含量的QTL
检测到与粗脂肪含量相关的QTL有9个,即Qcfc1~Qcfc9。其分布在连锁群LG1、LG3、LG4、LG7、LG8、LG10、LG11和LG13上,遗传贡献率为7.5%~27.2%,贡献率较高的(>15%)分别是位于LG7和LG8上160.0 cM和32.6 cM处的Qcfc4和Qcfc5,Qcfc4与SRAP标记m5e7-56共分离,Qcfc5与SSR标记xtxp230-386和SRAP标记m3e3-539紧密连锁。
2.2.3 茎叶比的QTL
检测到与茎叶比相关的QTL有12个,即Qslrw1~Qslrw12,其分别位于连锁群LG1、LG3、LG4、LG5、LG6、LG8、LG9、LG12、LG13和LG14上,遗传贡献率为11.6%~33.4%,超过20%的主效QTL有5个,其紧密连锁标记分别是m7e8-154、xtxp159-89、m10be7-957、m3e8-935和m7be7-184,贡献率最高的是位于LG1上 102.1 cM处的Qslrw1,对应标记为m7e8-154,作用方式表现为显性。
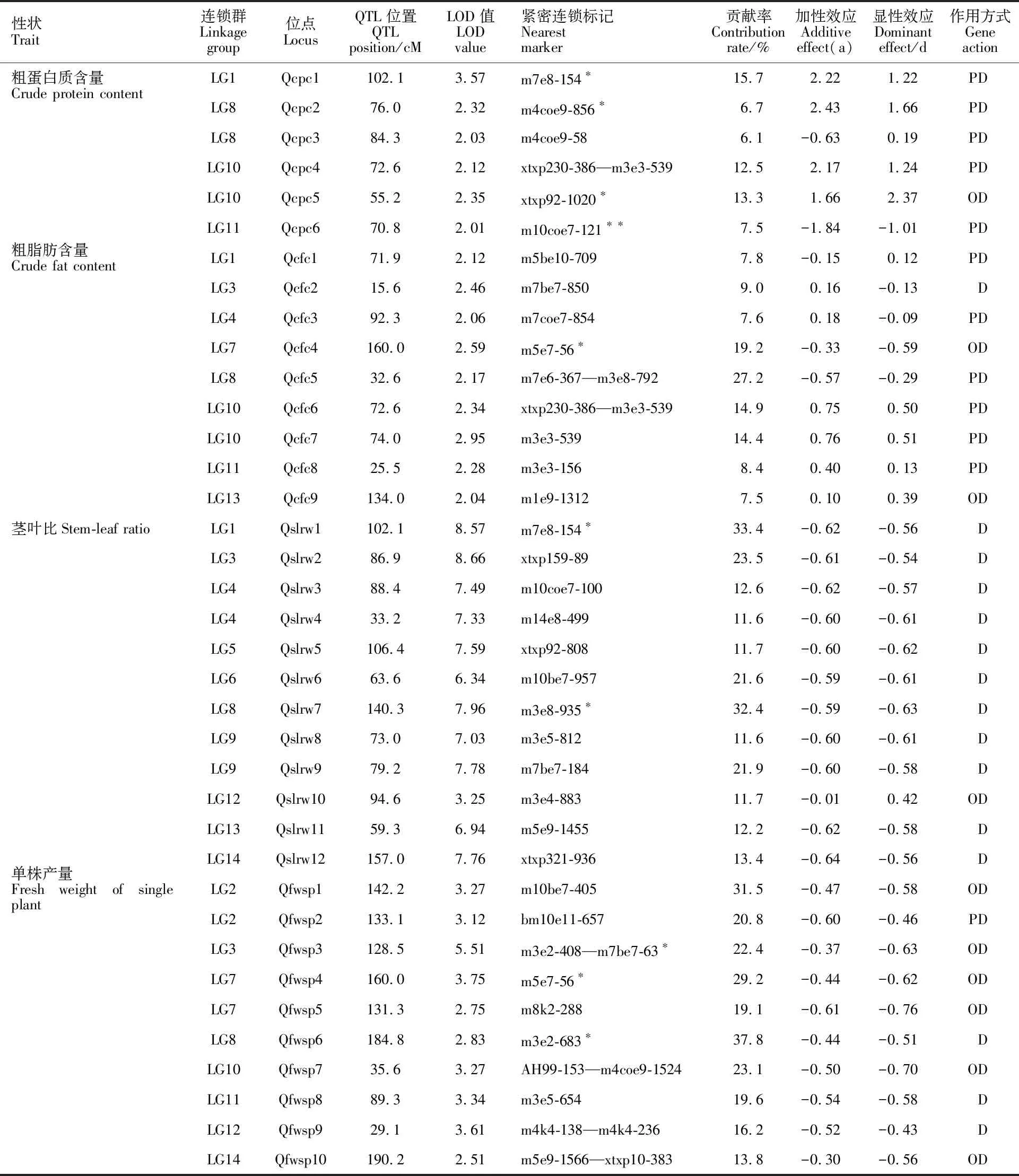
表2 冰草4个重要性状的QTL及其遗传效应Table 2 QTLs for four important traits and its genetic effect in wheatgrass
紧密连锁标记中,以英文字母m和bm开头的标记为SRAP标记,其余均为SSR标记。 PD:部分显性(|d/a|=0.20~0.80);D:显性(|d/a|=0.81~1.20);OD:超显性(|d/a|>1.20)。*和**:0.05和0.01水平偏分离标记。
SRAP markers are beginning with the letters of m or bm,and the rest are SSR markers. PD:Partial dominant(|d/a|=0.20- 0.80); D:Dominant(|d/a|=0.81-1.20); OD:Over dominant(|d/a|>1.20).* and **:Distorted markers of 0.05 and 0.01 levels.

(图续转下页 Be continued in next page)

图2 4个性状的QTLs在各连锁群上的分布
2.2.4 单株产量的QTL
检测到与单株鲜草产量相关的QTL位点有10个,即Qfwsp1~Qfwsp10。分别位于连锁群LG2、LG3、LG7、LG8、LG10、LG11、LG12和LG14上,遗传贡献率为13.8%~37.8%,超过20%的主效QTL 6个,其连锁标记分别是m10be7-405、bm10e11-657、m3e2-408—m7be7-63、m5e7-56、m3e2-683和AH99-153—m4coe9-1524。
3 讨 论
有研究表明,作物品质、产量、抗性等重要性状相关的QTLs存在着“一因多效”的遗传效应[22-24]。在牧草上,石 悦等[15]对高丹草产量相关性状的QTL研究发现,在同一连锁群上存在“一因多效”现象。本试验发现,控制冰草粗蛋白质含量和茎叶比的个别QTL在LG1上的102.1 cM处存在共位点标记m7e8-154,控制粗脂肪含量和单株产量的QTL在LG7的160.0 cM处存在共位点标记m5e7-56,控制粗蛋白含量和粗脂肪含量的QTL在LG10的72.6 cM处均与标记xtxp230-386—m3e3-539紧密连锁。进一步比较发现,这3个共位点标记QTL的遗传贡献率均较高,如控制粗蛋白质含量与茎叶比共位点标记m7e8-154的QTL遗传贡献率分别为15.7%和33.4%;控制粗脂肪含量和单株产量共位点标记m5e7-56的QTL遗传贡献率分别为19.2%和29.2%(表2)。这表明QTLs“一因多效”现象是植物的重要遗传效应之一,对这些“一因多效”的QTL进行跟踪和选择,可能有利于从正向遗传学角度开展目标性状QTL精细定位、基因图位克隆、功能解析和标记辅助育种的深入研究。
在植物性状QTL定位研究中,连锁系数LOD值的确定关系到QTLs在连锁群上的可信度。李慧慧等[25]研究指出,为尽可能多的定位到遗传效应较小的控制目标性状的QTLs,确保控制育种性状的基因不遗漏,需适度减小QTL阈值。Tanksley等[26]研究表明,高密度、高饱和度的分子遗传图谱适当降低检测QTLs时的LOD阈值,有利于提高定位的准确性。一般认为LOD阈值≥3.0时进行QTL位点检测效果较好,而当LOD<2.0时两位点间连锁效应较弱或不存在连锁;LOD>2.0时定位连锁群上的QTLs可以保证极小概率(P<0.05)错误的出现[27-28]。赵方媛等[29]以饲草型小黑麦F2群体的184个单株为材料,对其抗条锈病进行QTL定位分析时,将LOD≥2.0设为QTL阈值,共定位到6个抗条锈病QTLs。本试验在已构建出的四倍体杂交冰草分子遗传图谱上,用246个加倍植株F2群体分离单株为材料,定位冰草粗蛋白质含量等4个重要性状QTLs,由于该图谱为高密度图谱(含1 011个标记、平均间距2.19 cM),且各标记在连锁群上分布均匀、未出现聚集现象,因此确定LOD>2.0为阈值,使得同一性状检测到的QTL较多,保证了QTL检测结果的真实性和可靠性。
4 结 论
在前期已构建出的四倍体杂交冰草SRAP和SSR高密度分子遗传连锁图谱上,定位了控制粗蛋白质含量等4个重要性状的QTLs 37个,其中控制粗蛋白质含量性状的有6个、粗脂肪含量的有9个,茎叶比的有12个、单株产量的有10个。明确了各QTL在冰草14个连锁群上的分子标记位点和遗传效应,各QTL的遗传贡献率范围为6.1%~37.8%。
猜你喜欢
甘肃农业科技(2023年8期)2023-09-14 18:24:00
莫愁(2023年25期)2023-09-14 06:34:56
北方农业学报(2021年5期)2022-01-12 07:36:12
饲料工业(2017年15期)2017-01-08 09:31:18
畜牧兽医科技信息(2015年10期)2015-12-01 01:52:39
草原与草业(2015年4期)2015-03-18 07:22:18
当代畜禽养殖业(2014年8期)2014-03-22 07:51:45
动物营养学报(2013年12期)2013-09-20 03:05:48
农村百事通(2012年15期)2012-04-29 00:44:03
草食家畜(2012年2期)2012-03-20 13:22:39
