
野生茄子原生质体制备及电融合条件研究
2019-07-20 03:06郭欢欢陈钰辉杨锦坤刘富中
中国蔬菜 2019年7期
郭欢欢 陈钰辉杨锦坤 刘富中 张 映 连 勇
(中国农业科学院蔬菜花卉研究所,农业农村部园艺作物生物学与种质创制重点实验室,北京 100081)
原生质体是指去除细胞壁后被细胞膜包围的细胞,其应用研究主要通过原生质体融合即体细胞杂交创制新的植物种质。原生质体融合技术能够克服有性杂交中只有亲缘关系较近的物种间才能获得杂交后代的局限性,转移有性杂交方式中亲本不能转移的生物和非生物胁迫抗性,获得包括植物种间、属间及科间的体细胞杂种。原生质体融合技术已在茄属、烟草属、柑橘属、禾本科等植物中广泛开展(喻艳,2013;常胜合 等,2018)。
茄子栽培种和大多数茄子近缘野生种都属于茄科茄属一年生草本植物。自然界中,茄子栽培种一般抗逆性(生物逆境和非生物逆境)较差,而野生茄子表现出较好的抗逆性和抗病性(Rotino et al,2014),因此常用作砧木使用,常用的砧木材料为刺天茄(Solanum indicum)、水茄(S. torvum)及赤茄(S. mammosum)等。但由于近年来单一砧木品种使用年限较长及不同地区病害生理小种发生变异等原因导致砧木品种的抗病性下降,频繁出现了嫁接苗发生大面积死秧现象(王爽 等,2013),因此需要创制出新的砧木资源。由于不同野生茄子品种抗病种类及抗病能力不同(张红浩 等,2015),加上多数野生茄子间有性杂交不亲和(Daunay,2008),制约了砧木资源的改良。利用原生质体进行体细胞杂交可以克服有性生殖的障碍,将不同抗病种类和抗病能力的野生材料融合成体细胞杂种植株,创制出新的野生砧木 材料。
研究人员通过原生质体融合技术分别获得茄子栽培种和野生茄子S. sisymbriifolium、S. khasianum、 S. torvum、S. nigrum、S. aethiopicum 体 细胞杂交四倍体株系,并部分转移了野生资源中的优良抗病 性(Gleddie et al.,1986;Guri & Sink,1988;Sihachakr et al.,1988,1989;Daunay et al.,1993;Jarl et al.,1999)。前人虽已建立了茄子原生质体分离及融合的基本方法,但不同基因型原生质体游离所需酶类和酶解时间等条件存在差异,对不同融合亲本原生质体游离及融合条件仍需进一步探讨。因此,本试验以蒜芥茄(S. sisymbriifolium)和水茄(S. torvum)为融合亲本,主要探讨不同酶类浓度、酶解时间和电融合基本参数对原生质体游离及融合的影响,筛选出原生质体游离及电融合最佳条件,以期为亲本均为野生茄子的原生质体游离及融合提供试验基础,并为创制茄子砧木种质资源奠定 基础。
1 材料与方法
1.1 试验材料
水茄(S. torvum)和蒜芥茄(S. sisymbriifolium)种子由中国农业科学院蔬菜花卉研究所茄子课题组提供。2017 年12 月于茄科实验室组培室开始进行培养,原生质体游离材料为茄子试管苗嫩叶。
1.2 试验方法
1.2.1 外植体获得 挑取饱满健康种子,在无菌操作台中用70%的酒精浸泡30 s,10% NaClO3灭菌15 min,无菌水冲洗3 遍,吸干水分,然后接种于MS(3%蔗糖,0.75%琼脂)培养基中,培养瓶置于14 h 光照/10 h 黑暗、光照强度2 200 lx、温度25~27 ℃的组培室中培养。植株采用节间继代培养,原生质体游离使用的茄子苗在MS(2%蔗糖,0.75%琼脂)培养基中培养21 d,取上部嫩叶进行原生质体的游离纯化。
1.2.2 酶解液浓度对原生质体游离的影响 在无菌操作台中分别取水茄和蒜芥茄试管苗上部幼嫩真叶1~2 g,利用解剖刀将嫩叶切成1 mm 左右宽度,然后浸泡在酶液中,在(25±1)℃黑暗下酶解。纤维素酶(Onozuka R-10,Yakultv,Japan)设0.2%(m/V,下同)、0.3%、0.4% 3 个浓度梯度,离析酶(Macerozyme R-10,Yakultv,Japan) 设0.05%、0.1%、0.2% 3 个浓度梯度,9 个酶液组合,在不同酶液组合条件下对叶片原生质体游离情况进行显微镜观察比较,每个酶液组合重复3 次观测,选定最优酶解液。
1.2.3 酶解时间对原生质体游离的影响 选定最优酶解液后,测定不同酶解时间(9、10、11、12、13、14、15、16 h)对不同基因型茄子叶片原生质体产量的影响。每个酶解时间重复3 次,试验数据采用SAS 9.4 软件进行分析。通过比较不同酶解时间对原生质体产量的影响,确定最优酶解时间。
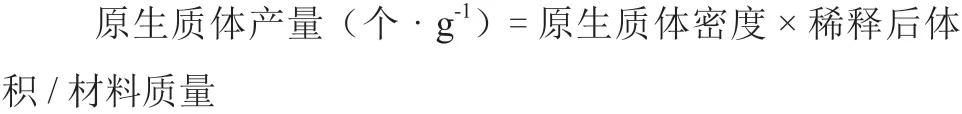
1.2.4 电融合参数对原生质体融合的影响 用电融合仪(ECM2001,USA)对游离纯化后的原生质体进行融合。对基本电融合参数进行设定,其中交流电场强度分别为60、70、80、90、100、 110 V · cm-1,交流电场作用时间以大多数细胞成串为3~4 个为准,直流脉冲强度分别为1 200、1 250、1 300、1 350 V · cm-1,直流电场作用时间分别为40、45、50、55 μs,每次试验在研究某一参数时,其他参数保持某个数值不变,通过显微观测细胞状态,确定最适电融合参数。每个处理重复3 次。
2 结果与分析
2.1 酶解液浓度对原生质体游离的影响
试验结果表明(表1),水茄和蒜芥茄叶片原生质体在0.3%纤维素酶+0.1%离析酶条件下镜检观测到的单细胞数量最多,其余酶解液浓度组合显微观察存在叶片解离不完全、单细胞量少、细胞破碎等情况,不利于原生质体的游离。显微镜下观测到在适宜的酶液浓度组合下,酶解充分,游离单细胞较多(图1-A),放大观测可见球体形状规则的、透亮的完整细胞(图1-B);酶液浓度不适易造成细胞破碎,显微镜下观测可见细胞呈不规则形状,有细胞碎片(图1-C、D)。
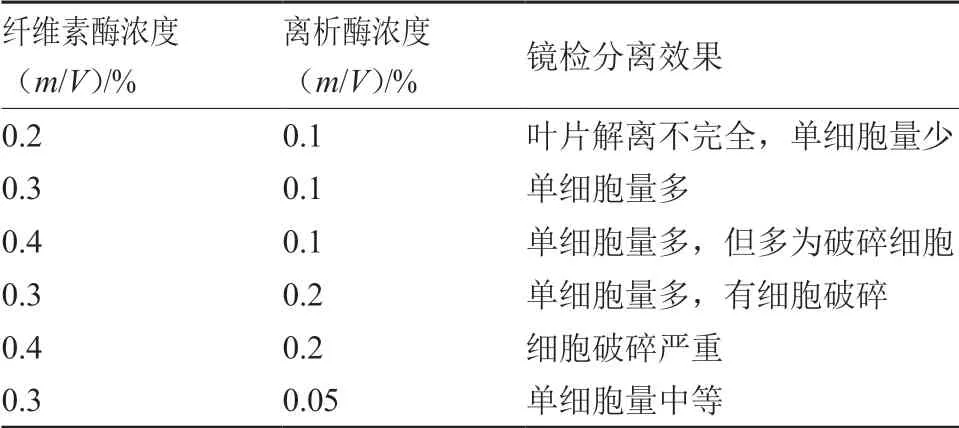
表1 部分酶解液组合下叶片原生质体游离效果观测
2.2 酶解时间对原生质体游离的影响
在最优酶解液浓度0.3%纤维素酶+0.1%离析酶条件下,对不同酶解时间下原生质体产量进行比较。结果表明(表2),蒜芥茄在13~15 h 酶解时间下的原生质体产量较高,其中14 h 达到最大值15.06×106个 · g-1。水茄在11~13 h 酶解时间下的原生质体产量较高,其中13 h 达到最大值15.92×106个 · g-1。酶解时间过短或过长都会影响原生质体产量,酶解时间过短,酶解不充分,不利于原生质体游离,影响原生质体产量;酶解时间过长,原生质体脱壁完全,导致质膜受到损伤,从而使原生质体死亡数量增加。
2.3 电融合参数对原生质体融合的影响
显微观测表明,在交变电场作用下,双亲原生质体沿电场方向形成很多平行排列的串珠,交变电场越大,串珠形成越快;交变电场作用时间越长,串珠形成也加长(图2-A)。然后施加直流脉冲电压后,串珠迅速解体,形成了很多长短不一的包括由两个原生质体形成的小串珠,部分小串珠很快融合在一起(图2-B、C),直流脉冲电压越大或作用时间越长,显微观测可见多核体多,原生质体破裂的多(图2-D、E)。本试验中确定基本电融合参数为交流电场强度80 V · cm-1,交流电场作用时间10 s,直流脉冲强度1 250 V · cm-1,直流电场作用时间 45 μs,次数1 次时,显微镜观测大多数细胞成串为3~4 个,大多数为两细胞之间发生融合,且细胞破碎少,融合效果好(图2-B、C)。

图1 野生茄子叶片原生质体游离效果
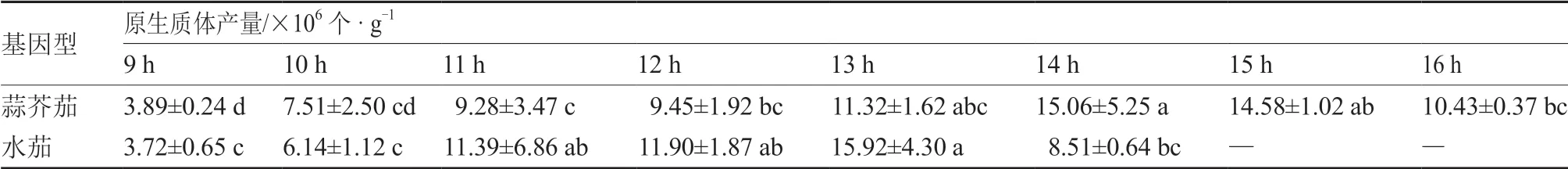
表2 酶解时间对不同基因型野生茄子叶片的原生质体产量的影响

图2 野生茄子原生质体在不同电融合参数条件下的融合效果
3 结论与讨论
酶的种类、浓度是影响原生质体游离的重要因素。纤维素酶用于在植物原生质体中裂解细胞壁,离析酶用于从叶肉细胞中分离出单个细胞。酶浓度过低,会使细胞壁裂解不充分,游离不出原生质体;酶浓度过高会损害原生质体,导致其破碎、活性下降。前人研究显示在喀西茄(S. khasianum)和癫茄(S. aculeatissimum)原生质体融合试验中,其叶片原生质体游离使用0.8%(m/V,下同)纤维素酶R-10 和0.2%离析酶R-10(Stattmann et al.,1994);栽 培 茄(S. melongena L.)分别 与 水茄 (S. torvum)和南美红茄(S. aethiopicum)进行原生质体融合时,其叶片原生质体游离使用1.5%纤 维 素 酶R-10 和0.5% 离 析 酶R-10(Sihachakr et al.,1989;Daunay et al.,1993);栽培茄和紫色非洲茄属(S. marginatum)叶片原生质体游离使用0.1%纤维素酶R-10 和0.2%离析酶R-10(Borgato et al.,2007)。不同基因型的植物由于叶片组织结构上的差异,从而使得在原生质体分离去壁酶解过程中酶解条件存在一定差异(喻艳,2013)。本试验中水茄和蒜芥茄叶片原生质体游离最适酶液浓度是0.3%纤维素酶R-10+0.1%离析酶R-10,在这个浓度下显微镜观测到的单细胞数量最多。由此可见,对于不同基因型植物,即使是同一属,其最适酶解浓度也不同,在进行原生质体游离时都需要研究确定最适酶解浓度。
酶解时间对植物原生质体产量有很大影响。酶解时间过短,不能达到较好的分离效果;时间过长,酶对已分离出的原生质体就会产生毒害作用。栽培茄(S. melongena L.)、水茄(S. torvum)和紫色非洲茄属(S. marginatum)的酶解时间为16 h (Guri & Sink,1988;Borgato et al.,2007)。本试验结果显示,蒜芥茄(S. sisymbriifolium)最适酶解时间是14 h,原生质体产量达最大值15.06×106个 · g-1,水茄(S. torvum)最适酶解时间是13 h,原生质体产量达最大值15.92×106个 · g-1。本试验结果和前人不同,说明不同基因型植物在相同酶种类和浓度条件下,获得最大原生质体产量所需要的酶解时间不同,选择合适的酶解时间才能获得大量高活性原生质体。
原生质体电融合可分解为两步,首先是交流电场使得原生质体3~4 个串在一起,然后给予直流脉冲使得串在一起的原生质体发生融合,交变电压过高,无论直流脉冲电压多么低,原生质体也会破裂,若交变电压过低,无论脉冲电压多么高,融合也很少发生,电融合参数影响融合效果(郭文武 等,1998;田国奎 等,2012)。不同基因型茄子原生质体电融合参数不同,研究表明,栽培茄(S. melongena L.)和水茄(S. torvum)或南美红茄(S. aethiopicum)使用电融合参数为交流电场强度125 V · cm-1,作用时间15 s,直流脉冲强度1 200 V · cm-1,作用时间20 μs,次数2 次(Sihachakr et al.,1989;Daunay et al.,1993); 栽 培 茄(S. melongena L.)和紫色非洲茄属(S. marginatum)使用电融合参数为交流电场强度125 V · cm-1,作用时间15 s,直流脉冲强度1 500~2 000 V · cm-1,作用时间50 μs,次数3 次(Borgato et al.,2007)。本试验中,交流电场强度为80 V · cm-1,作用时间10 s,直流脉冲强度1 250 V · cm-1,作用时间 45 μs,次数1 次时,显微镜观测到大多数细胞成串为3~4个,融合效果较好。与前人试验所用电融合参数相比,本试验所用电流强度和时间不同,说明不同基因型茄子在电融合时需要确定适当的电融合参数,参数的确定对能否获得大部分两细胞融合是非常必要的。
猜你喜欢
实用手外科杂志(2022年2期)2022-08-31
福建农业学报(2021年2期)2021-05-31
新农业(2020年21期)2020-11-19
科学导报·学术(2020年29期)2020-10-21
美与时代·美术学刊(2019年9期)2019-11-29
阅读与作文(小学高年级版)(2019年9期)2019-11-06
西南农业学报(2019年6期)2019-07-18
幸福家庭(2019年14期)2019-01-14
小天使·一年级语数英综合(2017年4期)2017-04-18
外语教学理论与实践(2014年2期)2014-06-21
