
叶形对银杏叶片质量-面积异速生长关系的影响
2019-07-19 06:43刘梦菂杨雅运时培建
安徽农业科学 2019年12期
关键词:叶面积
刘梦菂 杨雅运 时培建
摘要 [目的]探究银杏叶片质量和面积之间是否符合异速生长关系,并分析叶形对异速生长指数的影响。[方法]使用360片成熟的银杏叶片,根据长宽比的中位数将样品分为2组,使用不偏长轴法拟合叶片质量和面积的对数转换数据。[结果]发现银杏的叶片质量和面积之间存在异速生长关系,且异速生长指数大于1,表明叶片质量的增加与叶片面积的增加并不同步;同时还发现长宽比较小的叶片其异速生长指数的估计值更小,表明叶形特别是长宽比能显著地影响叶片质量和面积的异速生长关系。[结论]银杏叶片质量和面积的异速生长关系反映了叶片单位面积上干物质投资对叶片面积的收益递减规律,而宽叶片收益递减的效应较低,在一定程度上解释了银杏叶片中呈90°以上的扇形所占比例较大的生物学现象。
关键词 异速生长;叶面积;叶干重;不偏长轴法
中图分类号 S 792.95 文献标识码 A
文章编号 0517-6611(2019)12-0149-04
doi:10.3969/j.issn.0517-6611.2019.12.041
开放科学(资源服务)标识码(OSID):
Abstract [Objective]This study was performed to examine whether there is a scaling relationship between leaf mass and leaf surface area and to analyze the influence of leaf shape on the scaling exponent. [Method]360 mature leaves of Ginkgo biloba L. were collected from 10 healthy trees on the Nanjing Forestry University campus. We divided the leaves into two groups according to the quotient of leaf length to width (i.e., the broadleaved group vs. the narrowleaved group). Reduced majox axis was used to fit the logtransformed data of leaf mass vs. area for the pooled data and each group. [Result]The scaling relationship of leaf mass vs. area holds, and the estimate of the scaling exponent is greater than unity, which indicates that the increase of leaf mass does not keep with that of leaf surface area. We also found that the estimate of the scaling exponent of leaf mass vs. area for the broadleaved group was smaller than that of the narrowleaved group, which demonstrates that leaf shape that is represented by the quotient of leaf length to width has a significant impact on the scaling relationship between leaf mass and area. [Conclusion]The scaling relationship of leaf mass vs. area of G. biloba reflects the phenomenon of diminishing returns of the investment of leaf dry matter relative to the increase of leaf surface area. A lower extent of diminishing returns for the broadleaved group explained why the angles for most fanshaped leaves of G. biloba are greater than 90 degrees to some degree.
Key words Allometric growth;Leaf area;Leaf dry weight;Reduced majox axis
叶片是植物进行光合作用、蒸腾作用以及维系自身热平衡的主要器官,是其对全球气候变化最为敏感的地上部分[1-2]。而且植物大部分的碳通量是由叶片表面积决定的,所以叶片生物量的分配能够影响植物很多重要的表型特征,比如叶柄和叶脉生物量的分配限制了叶片的大小和数量[3]。因此,不同生境下叶片质量和面积之间的异速生长关系已经成为许多学者探索的主题[4-5]。
叶片质量(LM,g)与面积(LA,cm2)之间的关系通常使用幂函数来描述:LM=αLAβ[6],其中α和β是待拟合常数,β被称为异速生长指数。当β=1时,叶片质量和面积的增加是同步的,这种比例关系不受环境等因素差异的影响[7];当β>1时,叶片质量和面积的增加并不同步,即叶片质量的增量要大于叶片面积的增量,这即Niklas等[8]提出的“收益递减假说”。异速生长理论在动物学研究中首先得到了应用[9],但它也可以帮助解释植物群落内的叶片大小和频率分布。例如Toledohernández等[10]利用在狮子鱼入侵范围内采集的17个已公布的长度-重量关系,發现异速生长参数存在显著的区域差异,他们观察到的空间模式与其他关注遗传学或年龄的研究者的结果一致。Enquist[11]利用异速生长关系的一般理论,预测了维管植物的比例和植物群落的特征是如何随着植物大小变化而变化的,为异速生长提供了新的证据,同时他指出在叶木质部的解剖学和生理学属性中没有这种异速生长关系。Niklas等[8]研究了叶面积与叶干重、水分和N/P之间的比例关系,虽然它们在统计上存在差异,但这些变量之间的关系在6个种群和19个独立物种之间的数值是相似的。在众多的异速生长指数中至少有1个不等于1,这说明存在1个对叶性状有抑制影响的异速生长指数,它反映了所有植物的一个基本特征。Pan等[12]研究了海拔对质量-面积异速生长指数的影响,发现在低海拔处,异速生长指数在1附近,而高海拔处该指数小于1。姚靖等[13]和王文娟等[14]分别研究了五角枫[Acer pictum subsp.mono(Maxim.)Ohashi]和胡杨(Populus euphratica Oliv.)叶性状的异速生长随发育阶段的变化,发现不同发育阶段叶性状的异速生长指数并不相同。研究异速生长关系的指标一般有叶面积、叶干重、水分含量、叶体积、叶柄干重、叶脉密度、N/P等[15-16],该研究主要关注叶片质量和面积之间的关系。
目前对被子植物在物种多样性和生态学上的研究较为丰富,而对裸子植物的研究比例相对较小。银杏(Ginkgo biloba L.)是从裸子植物的祖先进化而来的,它提供了与开花植物的大数据集进行有用的“外群”比较,对于研究植物进化史具有重要的理论意义。近年来对于银杏叶片的研究大多集中在叶片提取物的生理、生化研究等方面[17-19],对叶片性状的异速生长关系研究相对较少,Christianson等[20]按照雌雄、长短枝等性状将银杏叶片划分成了7个描述叶质量和面积的统计学模型,发现这7种比例与利用SLA(specific leaf area)值确定的叶片类别之间并没有一致性,银杏SLA值的总体范围很大,反映了冠层结构叶片与一些生态现象指标的相关性。Niinemets等[21]研究了叶片形状和叶脉模式对温带植物叶片内用于支撑叶片物理性状生物量投资的影响,发现心形和卵形叶片对于机械支撑的要求明显较少,因为它们的叶片质量靠近叶的基部,而椭圆形或倒卵形叶的叶片质量位于离叶基部较远的位置。但是相关研究并没有对银杏独特的扇形叶片进行比较,它的叶内都是细脉,没有明显的主脉,但基本上跨越叶片的整个长度,保持彼此相等的距离。笔者以银杏为材料,分析了其成熟叶片质量和面积的异速生长关系,并探讨了叶形对叶片质量-面积异速生长指数(反映了植物光合能力的强弱)的影响。了解叶片大小和功能性状之间的关系将提高人们对叶片如何保持碳平衡和影响整株植物适应性的理解,这对于建立叶片生长的生理生态模型具有重要的价值。
1 材料与方法
1.1 材料
2018年11月28日在江苏省南京市南京林业大学校园内(118°48′52″ E,32°03′09″ N)采集叶片,选取生长健康的银杏成树10棵(胸径>5 cm,树高>5 m),在树冠外围东、西、南、北4个方向采集36片完全展开且没有病虫害的叶片,一共360片。将采集到的叶片立即放入塑料自封袋(28 cm×20 cm)内运输,为防止叶片失水皱缩,采完后立即回实验室进行指标测定。
1.2 设备
Aficio MP7502扫描仪(Rocoh公司,日本东京);XMTD-8222烘箱(上海精宏实验设备有限公司,中国上海);ME204-02电子天平(精度0.000 1 g,梅特勒-托利多仪器(上海)有限公司,中国上海)。
1.3 方法
首先将叶子擦拭干净,并剪去叶柄。然后用扫描仪进行叶片扫描,统一保存为分辨率400 dpi的位图(Bitmap),随即装在纸质信封(33 cm×24 cm)放入烘箱,先在105 ℃的环境下进行2 h的杀青,然后在80 ℃的环境下烘干48 h至恒重,取出冷却至室温后用电子天平称重记录。扫描的叶片图像用PS(Adobe Photoshop CC)处理得到叶片边缘的黑白位图,通过Matlab(2009a)和R软件(3.2.2)计算叶片的长度、宽度和面积[22]。该研究将银杏叶片左右两端的最长处定义为宽,从叶基处引一条垂直于宽的线段定义为长(图1)。
1.4 数据分析
为了稳定叶片质量和面积的方差,对叶片质量和面积的幂函数方程两边同时取对数,得到如下公式:
y=γ+βx(1)
式中,y=lnLM,γ=lnα,x=lnLA。为了计算上述方程的斜率和截距,这里使用不偏长轴法估计模型参数,因为当y和x变量之间没有明显的先验函数关系时,以及当x变量受到测量误差时,不偏长轴回归更合适,数据分析与做图均使用R软件完成。
2 结果与分析
2.1 叶片质量和面积的异速生长关系分析
图2展示了对银杏叶片质量和面积对数转置数据的线性拟合结果,横轴表示叶片面积的自然对数,纵轴表示叶片质量的自然对数,可以看出两者呈现出较好的线性关系(R2=0.865),其中斜率为1.549,β>1,这表明成熟叶片质量的增量往往比面积的增量要大,即两者是非同速增长的。换言之单位面积上的叶面积干物质的投入随着叶面积的增大而增大,对应单位叶面积构建和维护成本的增加。
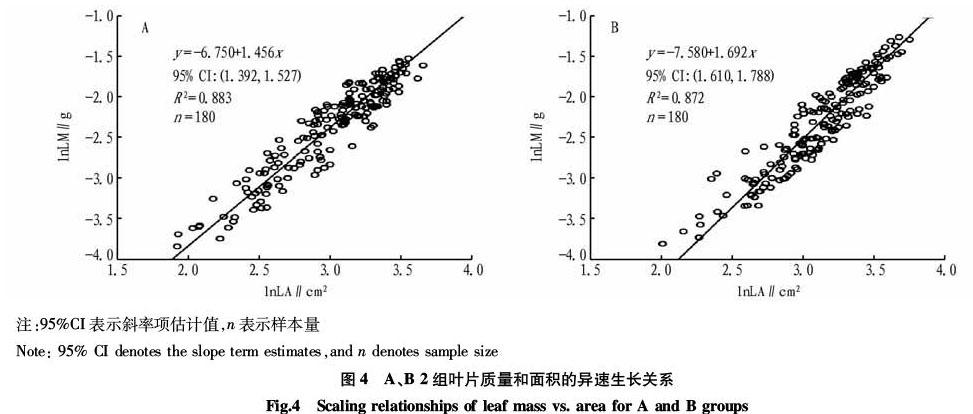
2.2 叶形对叶片质量-面积异速生长关系的影响
该研究中银杏叶片的长宽比为0.46~0.85,这360 片叶片长宽比的中位数是0.6。笔者将长宽比为0.46~0.60的叶片设为组A,长宽比>0.60~0.85的叶片设为组B(图3),分别对每组
叶片质量和面积的自然對数数据做散点分布图(图4)。并使用不偏长轴法拟合2组数据,发现2组内叶片质量和面积均呈较好的线性关系(R2分别为0.883和0.872),且组A的斜率小于组B的斜率。这说明长宽比较小的叶片,其叶片质量随叶面积变化的效应更小。2组数据线性拟合的斜率项95%的置信区间不重合,表明2个斜率存在显著性差异。
3 讨论
3.1 叶片质量和面积异速生长关系的成因探讨
通常植物可利用的环境资源是有限的,当对某一功能性状的投资较多时,植物必然会减少对其他性状的投资[23]。Navas等[24]和Whitman等[25]通过对大量植物性状特别是草本植物的研究,提出植物要达到自身的“平衡”“生存”和“生长繁殖”三大目的,就需要在功能性状之间进行资源的优化配置,调整、转换或者补偿自身功能,这些分配和均衡最终会表现在宏观的植物器官的组织结构以及生理性状上。该研究表明成年银杏叶片质量和面积之间是符合异速生长关系的,这与姚靖等[13]和杨琼等[26]的研究结果一致,同时他们分别研究了在不同发育阶段五角枫、胡杨叶片质量和面积的关系,提出这种异速生长关系随着发育阶段的不同而变化。植物在幼苗和幼树阶段,叶片质量和面积的生长比例指数在1附近,表现为同速增长的关系;在成树阶段,叶片质量和面积的异速生长指数大于1,这表明对于较大的叶片,植物选择投资更多的生物量用于构建叶片的机械结构。因为在这个阶段成熟叶片的结构相对稳固,叶片内支持组织的含量更高,使得内部物质的扩散效率降低,植物会相对地增加对叶片机械组织(例如叶柄和叶脉)的生物量投资,从而能将水分更快地运输到叶肉细胞[27]。同时随着树木的增高和叶片的逐渐变大,叶片所受的风力、水力限制也在增加,需要更牢固的机械结构去支撑叶片。杨琼等[26]通过对不同发育阶段的胡杨叶片进行研究,提出植物为了避免叶片的相互遮挡,将更多的能量用于叶柄和叶脉的构建。Sun等[28]对生长在中国东南武夷山不同海拨的5种竹种进行了研究,发现其叶片面积和干重的异速生长关系对海拔和光照并不敏感,但Pan等[12]提出叶片面积和干重的异速生长指数随海拔升高而增大。诸如气候、光照、海拔等因素虽然对异速生长有影响,但改变的只是异速生长指数数值的大小,暗示了叶片质量和面积的异速生长关系是普遍存在的,环境因子可能只改变这种异速生长的程度[29]。
3.2 葉形对异速生长影响的物理解释探讨
功能和支持结构投资之间的权衡是叶片生理功能活动的主要决定因素,叶形和叶脉的分布模式与生物量的投资密切相关[30]。叶片的长宽比决定了整个叶片的结构形状[14],该研究发现成熟银杏叶片质量和面积存在异速生长关系,特别是长宽比较小的叶片其异速生长效应更小,这表明改变支持叶片生物量和叶片大小关系的一个因素可能是叶形。叶片质量是从叶基往顶点分布的,其中大多数叶片质量靠近叶片基部,这决定了总体弯曲力矩和公共区域的质量与叶片所需的支撑[21]。长宽比小的叶片实际上具有较短的杠杆臂,那么叶片中用于自我支撑的生物量投资就会减少,对于叶片面积的生物量投资就会增加,从而使得叶面积和叶干重的异速生长效应更小。宽叶片的收益递减程度较低,这在一定程度上解释了为什么银杏叶片中呈90°以上的扇形占的比例比较大[31]。
3.3 收益递减的成因探讨
SLA代表了投入的潜在回报率,这个系数是成熟叶片的一个特征,已被证明与植物的生长速率有关,其在不同类群中是变化的[29]。该研究中银杏叶片质量和面积的增长并不同步,随着叶面积的增加,SLA是逐渐减小的,即叶片面积增加所获得的收益并不一定能弥补叶片支撑成本的增加,表明银杏叶片符合“收益递减假说”[8]。一般而言,较大的叶片有更多的接触面积以拦截光线,这无疑会增强光合作用。但相应的大叶片比小叶片需要更多的生物量投资用于构建复杂的叶脉和叶柄等结构,以保证养分和水分的有效运输[32]。从叶片自身角度分析,并不是叶片越大对植物越有利,叶片较大的物种并不比叶片小的物种有更多的优势。叶水平上的生物量分配策略也是植物叶片大小优化的内在因素之一,这种现象揭示了在个体物种水平和功能物种群体水平上对成熟叶片大小的一个限制因素,使其不能无限增大[8]。但有一些学者认为这种“收益递减”的现象并没有在本质上限制叶片的大小,只是提供了一种补偿机制。这些比例关系在物理上是不可避免的,例如作为叶片新陈代谢中产生的惰性物质的积累导致了叶片干物质的增加,并不是植物权衡机制的作用[33]。
通常单位面积的叶片质量代表了光合能力,这实际上暗示了叶片水分(叶片鲜重减去叶片干重)与叶片干重成正比的假设[29]。然而Huang等[34]基于对15种阔叶树种叶片干重和鲜重的研究,发现叶片干重与鲜重之间存在异速生长关系,这就意味着叶片水分量随着叶片干重的增加而不成比例的增加。他们还发现无论从个体物种层面还是按科汇总数据层面,使用叶片鲜重和面积计算的异速生长指数都低于使用叶片干重和面积计算的异速生长指数,且叶片鲜重与面积异速生长关系的拟合优度要优于叶干重与面积的关系,这表明叶片鲜重可能比叶片干重更能反映与光合作用和呼吸相关的叶片的生理功能。他们建议在考察叶片生物量与面积的异速生长关系时宜使用叶片鲜重来表示叶片生物量,以避免出现高估“递减收益”效应的情况。
4 结论
该研究发现成熟的银杏叶片质量和面积存在异速生长关系,且其异速生长指数的估计值大于1,这说明叶片面积的增量低于叶片干物质的增量,即叶片面积增加所获得的收益并不一定能弥补叶片支撑成本的增加。此外,发现叶形(以长宽比表示)对异速生长指数具有显著影响,长宽比较小的银杏叶片(即夹角越大)其异速生长指数的估计值较小,那么叶片面积的收益递减趋势也越小,这在一定程度上也解释了为什么银杏树上宽叶片(夹角大于90°)较多的现象。
参考文献
[1] 李曼.武夷山主要森林类型植物根、茎、叶功能性状关联研究[D].福州:福建师范大学,2017.
[2] WRIGHT I R,GROOM P K,LAMONT B B,et al.Leaf trait relationships in Australian plant species[J].Functional plant biology,2004,31(5):551-558.
[3] NIINEMETS ,PORTSMUTH A,TOBIAS M.Leaf size modifies support biomass distribution among stems,petioles and midribs in temperate plants[J].New phytologist,2006,171(1):91-104.
[4] KENZO T,TANAKAODA A,MASTUURA Y,et al.Morphological and physicochemical traits of leaves of different lifeforms of various broadleaf woody plants in interior Alaska[J].Canadian journal of forest research,2016,46(12):1475-1482.
[5] VASSEUR F,VIOLLE C,ENQUIST B J,et al.A common genetic basis to the origin of the leaf economics spectrum and metabolic scaling allometry[J].Ecology letters,2012,15(10):1149-1157.
[6] POORTER H,DE JONG R.A comparison of specific leaf area,chemical composition and leaf construction costs of field plants from 15 habitats differing in productivity[J].New phytologist,1999,143(1):163-176.
[21] NIINEMETS ,PORTSMUTH A,TOBIAS M.Leaf shape and venation pattern alter the support investments within leaf lamina in temperate species:A neglected source of leaf physiological differentiation?[J].Functional ecology,2007,21(1):28-40.
[22] SHI P J,RATKOWSKY D A,LI Y,et al.A general leaf area geometric formula exists for plantsevidence from the simplified gielis equation[J].Forests,2018,9(11):1-15.
[23] ZHANG D Y.Plant lifehistory evolution and reproductive ecology[M].Beijing:Science Press,2004.
[24] NAVAS M L,ROUMET C,BELLMANN A,et al.Suites of plant traits in species from different stages of a Mediterranean secondary succession[J].Plant biology,2010,12(1):183-196.
[25] WHITMAN T,AARSSEN L W.The leaf size/number tradeoff in herbaceous angiosperms[J].Journal of plant ecology,2009,3(1):49-58.
[26] 楊琼,李征珍,傅强,等.胡杨(Populus euphratica)叶异速生长随发育的变化[J].中国沙漠,2016,36(3):659-665.
[27] POORTER H,EVANS J R.Photosynthetic nitrogenuse efficiency of species that differ inherently in specific leaf area[J].Oecologia,1998,116(1/2):26-37.
[28] SUN J,FAN R,NIKLAS K J,et al."Diminishing returns" in the scaling of leaf area vs.dry mass in Wuyi Mountain bamboos,Southeast China[J].American journal of botany,2017,104(7):993-998.
[29]MILLA R,REICH P B.The scaling of leaf area and mass:The cost of light interception increases with leaf size[J].Proceedings of the royal society B:Biological sciences,2007,274(1622):2109-2114.
[30] GODIN C,SINOQUET H.Functionalstructural plant modelling[J].New phytologist,2005,166(3):705-708.
[31] SHI P J,LIU M D,YU X J,et al.Proportional relationship between leaf area and the product of leaf length and width of four types of special leaf shapes[J].Forests,2019,10(2):1-13.
[32] RUNIONS A,FUHRER M,LANE B,et al.Modeling and visualization of leaf venation patterns[J].ACM Transactions on Graphics,2005,24(3):702-711.
[33] WITKOWSKI E T F,LAMONT B B.Leaf specific mass confounds leaf density and thickness[J].Oecologia,1991,88(4):486-493.
[34] HUANG W W,RATKOWSKY D A,HUI C,et al.Leaf fresh weight versus dry weight:Which is better for describing the scaling relationship between leaf biomass and leaf area for broadleaved plants?[J].Forests,2019,10(3):1-19.
猜你喜欢
农学学报(2022年8期)2022-11-16
农业科技通讯(2022年2期)2022-02-28
浙江农业科学(2022年2期)2022-02-25
北京园林(2021年2期)2022-01-19
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02
新疆农业科学(2020年1期)2020-02-14
亚热带植物科学(2019年2期)2019-07-22
新疆农垦科技(2018年10期)2018-04-12
甘肃林业科技(2016年4期)2016-05-11
中国麻业科学(2015年5期)2015-12-28
