
西藏高寒退化草地短期恢复后的抗牧性能研究
2019-07-15 09:26徐雅梅仝淑萍关法春苗彦军余成群蔡翠萍
西南民族大学学报(自然科学版) 2019年3期
徐雅梅,仝淑萍,关法春,苗彦军,余成群,蔡翠萍
(1. 西藏农牧学院,西藏 林芝860000;2.吉林省农业科学院,吉林 长春 130033;3.中国科学院地理科学与资源研究所,北京 100101;4. 伊犁职业技术学院,新疆 伊宁 835002)
青藏高原地区属于生态脆弱区[1-2],过度放牧等人为因素直接影响植物群落结构和组成[3-5],并对土壤质量产生不利影响,进而导致草地系统植被、土壤状况的整体退[6-7]. 鉴于草地退化对生态环境保护和农牧业生产造成的不利影响[2],西藏广泛应用牧草直播、牧草移栽、围栏封育生态等恢复措施[8],开展退化草地恢复工作.
过度放牧是西藏高原退化草地恢复过程中经常发生的生产行为之一,其行为会引起草地植物群落物种组成、系统多样性和生产力等发生变化[2-9],因而可以用上述指标来评估草地的抗牧性能.当退化草地恢复到一定程度后,如果再次进行放牧,草地群落植物种类组成、多样性特征及生产力等将如何变化? 植物群落抵御再次放牧的能力如何? 目前对此研究涉及有限.为此,本研究通过对比试验,研究牧草移栽措施恢复三年后,已恢复草地再次遭到过度放牧后的草地生产力和生物多样性状况,以期分析和明确西藏高寒地区退化草地短期恢复后的抗牧能力,从而为今后高原退化草地生态恢复策略的制定提供现实依据.
1 试验材料与方法
1.1 试验地背景
试验地位于西藏中部达孜县中科院拉萨高原生态试验站内(N:29°41′,E:91°20′),海拔高度约3 680 m,属高原温带半干旱季风气候区,年平均气温约7.5 ℃,年降水量约430 mm,80%以上的的降水集中在6 ~9 月.试验地土地沙砾化严重,植被稀疏,主要以白草(Pennisetum flaccidum)为主,草地植被退化严重.
1.2 试验设计
2012 年春季选择退化程度均一、地势平坦无起伏的退化草地,并进行连续观测. 试验设置移栽处理(Transplant),以相邻天然草地为对照(CK),小区面积为15 m×10 m,重复3 次.2012 年6 月末选取9 ~11 叶龄、长势一致的紫花苜蓿与草木樨营养钵苗,按照4:1 比例移栽至试验地内,移栽株行距70 cm ×30 cm、深度5 ~6 cm,试验期间无牲畜进入;在2012 ~2014 年期间,除了每年秋季上冻前和春季牧草返青时漫灌2 次外,所有样地不再进行任何管理.
移栽处理样地进行围封3 年后,2015 年春季起由当地村民进行自由放牧活动(主要为放牧动物为牛),草地不再进行任何管理.
1.3 测试项目与方法
1.3.1 数据采集
根据草食家畜采食习性,草地植物群落分为3 个功能群:豆科(Legumes)为可食豆科物种,但不包括毒草;禾草(Grasses)为可食禾草物种;杂类草(Forbs)为除以上功能群之外的物种.2015 年8 月中旬开始植被采集,在不同处理小区内随机设置1 m ×1 m 的小样方,3 次重复,记录样方内的植物种类、植株株高(精确到1mm),并采用方格法测定植被盖度,最后将样方内各种植物齐地面剪下,分种装袋带回实验室105 ℃杀青30 min,80 ℃下烘干至恒重,计算地上生物量.
1.3.2 测试项目
计算样地植物种类重要值(IV),生物多样性指标包括Berger-Parker 多度(Pi)、Shannon-Wiener 多样性指数(H′)、Pielou 均匀度指数(E)、Simpson 优势度指数(D)和Margalef 物种丰富度指数(DMG).具体如下:
IV =(相对高度+相对频度+相对密度+相对盖度+相对生物量)/5,Pi =ni/N,

式中:ni为样地某物种的重要值,N 为物种的总重要值,S 为样方植物群落总物种数.
采用Excel 2003、SPSS 21.0 软件处理数据,t-检验法、方差分析法进行差异显著性分析.
2 结果与分析
2.1 草地植物物种和密度
表1 所示是移栽处理和对照放牧后草地植物的物种及密度情况,共采集到植物6 种,豆科1 种、禾草3 种、杂类草2 种.其中移栽处理有5 种植物,包括豆科1 种、禾草2 种和2 种杂类草,对照有4 种植物,包括禾草2 种和杂类草2 种. 在移栽处理中,其优势种为白草,密度为363.56 株/m2,而移栽紫花苜蓿的密度仅为8.89 株/m2,在对照中,其优势种也是白草,密度为388.89 株/m2,处理间差异不显著(P >0.05;n=9).

表1 放牧后植物的物种密度Table 1 Species density of plants after grassland grazing
2.2 草地植物功能群密度、盖度
移栽处理和对照草地放牧后的各功能群密度和盖度情况如表2 所示.移栽处理豆科植物的密度降至8.89 株/m2,盖度降至1.33%l 移栽处理禾草密度略低于对照,但盖度高于对照,处理间差异不显著(P >0.05;n =9). 移栽处理杂类草密度与盖度均低于对照,处理间差异显著(P <0.05;n =9).而移栽处理的总密度为405.78 株/m2,为对照的74.35%,移栽处理的总盖度为39.56%,是对照的1.41 倍,上述指标处理间差异均不显著(P >0.05;n =9).

表2 移栽处理和对照三个功能群的总密度、总盖度Table 2 Weed species and density of three functional groups under different treatments
2.3 草地植物功能群生物量
表3 是移栽处理和对照草地各功能群的地上生物量情况.移栽处理豆科地上生物量仅为0.41 g/m2,对照为零;移栽处理禾草生物量为11.21 g/m2,高于对照(6.36 g/m2),而杂类草生物量为0.46 g/m2,低于对照的0.87 g/m2,但上述指标处理间差异不显著(P >0.05;n =9).移栽处理总生物量12.08 g/m2,为对照的1.67 倍,处理间差异不显著(P >0.05;n =9).
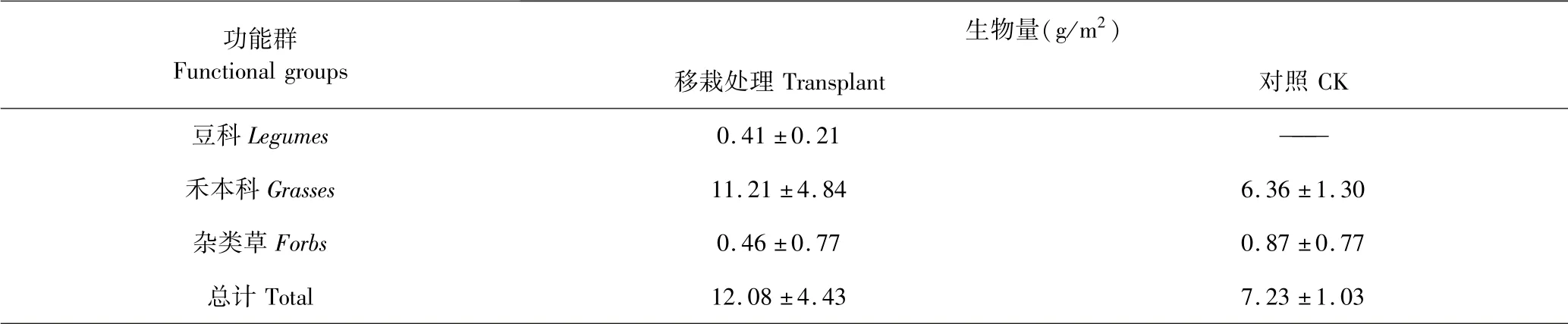
表3 草地各功能群植物地上生物量Table 3 Aboveground total biomass of functional groups biomass under different treatments
2.4 草地生物多样性指数
多样性指数的数据情况如表4 所示.移栽处理的Shannon-Wiener 多样性指数是对照的1.03 倍,Pielou均匀度指数是对照的85.33%,Margalef 物种丰富度指数是对照的1.33 倍,Simpaon 多样性指数是对照的1.03 倍,上述指标处理间差异不显著(P >0.05;n =9).

表4 植物群落多样性指数Table 4 Biodiversity indices of the plant community
3 讨论
草畜家畜的采食会直接诱导草地植物群落发生变化[10],如晚期演替的优势种被早期或中期演替的优势种所替代[11],同时在结构上往往出现高大的禾草被中等或矮生禾草取代[12]. 试验样地经过前期处理后,除了苜蓿和草木樨为人工引入的植物种类外,处理间仅在马唐(var esculenta)、黑穗画眉草( Eragrostis nigra.)有所差异,这可能与试验地地表完全沙砾化的立地条件,导致地上生长的植物种类稀少有关;不过,放牧对植物密度等指标影响相对于种类更大,移栽处理植物总密度及各功能群植物密度均明显低于对照,草食家畜偏好性采食行为[13],可能是导致有苜蓿存在的移栽处理样地植物密度低于对照的主要原因.
牧草移栽恢复措施的后续效应在放牧后的移栽处理样地上也有所体现,虽然移栽处理样地植物密度低于对照,但在采样过程中发现移栽处理样地的植物更为粗壮,尤其是白草、马唐匍匐生长较多,这与以往由于牲畜经常采食、践踏引起的禾草匍匐生长结论一致[14].由此导致移栽处理样地植物地上生物量和盖度均高于对照,同时提高了群落物种多样性,但群落均匀度指数却有所降低,这可认为是放牧因素和人为因素(移栽措施)共同作用的结果.
以往在本研究的试验样地上曾进行的退化草地人工移栽恢复措施,显著地提高了地上之植物生物量和生物多样性相关指标,实现了退化草地的短期快速恢复与植被重建[15],促进了生境条件的改善和系统生产力的提高[15-16]. 但短期恢复后的试验样地再度放牧后,其草地生产力和多样性指标与对照相比,差异并不大,说明短期恢复的退化草地,其抗牧能力有限.
4 结论
移栽与围栏封育相结合的措施恢复的退化草地在再次遭遇过度放牧侵害的情况下保留了比单纯围栏封育恢复草地更多的物种,并且而移栽处理的总密度为对照的74.35%,总盖度为对照的1.41 倍,总生物量为对照的1.67 倍,差异均不显著(P >0.05;n =9).Shannon-Wiener 多样性指数是对照的1.03 倍,Pielou 均匀度指数是对照的85.33%,Margalef 物种丰富度指数是对照的1.33 倍,Simpaon 多样性指数是对照的1.03 倍,均无明显差异(P >0.05;n =9). 以上数据均表明短期的移栽封育恢复措施尽管起到了提高草地抗牧性的作用,但并不能显著提高恢复草地抗牧性.因此退化草地经移栽封育短期恢复后,其抗牧能力并没有改观,高寒地区的重度退化草地应采取长期的恢复措施.
猜你喜欢
林业资源管理(2022年6期)2023-01-18
现代园艺(2021年23期)2021-12-01
新农业(2020年18期)2021-01-07
花卉(2020年24期)2021-01-04
黑龙江工程学院学报(2020年5期)2020-10-21
水土保持研究(2019年6期)2019-10-19
畜牧兽医科学(2019年24期)2019-09-10
南方农业·下旬(2017年11期)2018-01-09
绿色科技(2017年15期)2017-09-01
中国饲料(2017年15期)2017-08-22
