
碳、氮稳定同位素在构建海洋食物网及生态系统群落结构中的研究进展
2019-07-15 07:53张硕高世科于雯雯李士虎
水产养殖 2019年7期
张硕 ,高世科,于雯雯 ,李士虎
(1.上海海洋大学海洋科学学院,上海 201306;2.江苏省海洋水产研究所,江苏 南通 226007;3.淮海工学院海洋生命与水产学院,江苏 连云港 222005)
通过生态系统中的能量传递,生物之间形成了一种错综复杂的关联性,并在彼此之间构成直接、间接的网络结构,称为食物网[1]。相对于陆地生态系统,海域生态系统中的食物网结构常被用于连接海陆生态系统间的能量流动和物质转化,是生态学研究的一个重要领域[2,3],而群落结构和消费者营养级的确定是其不可分割的一部分[4]。传统海洋生物的食性研究是通过分析胃肠道中的食物组成及数量来确定其营养级[5],或者通过生物标志化合物分析法等[6],但这些方法均存在较大局限性,无法准确体现不同消费者的营养级,成为阻碍该领域发展的重大障碍。
近年来,稳定同位素技术被用于分析生态系统中的能量流动和物质交替变化[7-8],通过在食物链中富集度较低的碳(δ13C)稳定同位素比值来追溯捕食者食物的来源,利用氮(δ15N)稳定同位素在营养级间相对稳定的富集度来计算生物的营养级[9]。稳定同位素技术的优点在于克服了以往方法的复杂性和单一性[10],对了解生态系统中元素循环、物质能量循环提供了极大帮助。在海洋生态系统中,碳(δ13C)、氮(δ15N)稳定同位素主要用于构建食物网、确定营养级以及分析生物的食性来源,综合考虑了初级生产者到顶级消费者在食物网中的营养级时空变化及各层级消费者的食性特征及作用[11]。
Urey等[13-14]在1936年首次用化学交换法富集了13C,15N和34S等稳定同位素,于1947年发表了热力学相关文章,为现代同位素地球化学提供了理论基础;至20世纪50年代后期,科学技术的进步和更新使得稳定同位素技术得到迅速发展[15,16];1987年,Peterson等[10]发表了一篇稳定同位素在生态系统中的研究报告,引起学术界的极大关注;2002年,Phillips等[16]提出IsoSource模型在生态环境中的应用,给出了同位素定源贡献的范围。此后该模型被广泛应用于解决生态系统食物网的食物来源和营养级等问题层面。我国针对稳定同位素技术的研究起步稍晚,20世纪80年代以来,一系列大型国际涉海计划的实施使得稳定同位素在海洋生态系统的研究日益增多[18-19]。1999年,蔡德陵等人[20-21]首次基于稳定同位素技术构建崂山湾海洋生态系统的食物网结构,此后,我国各大海域、港湾及渔场的食物网基础构建及营养级确定成为重要研究对象[22-24]。可见,稳定同位素已然成为构建海域生态系统食物网、探究营养级及其动态关系的一种重要手段。
1 稳定同位素技术在食物网结构和生物营养级方面的应用
作为食物网基础研究的重要指标之一,营养级不仅反映物种群落结构的变化,还是用于海洋生态评价及效果评估的重要依据[24]。因此,在海洋生态系统中,不同生物营养级的确定及物质循环、能量传递对食物网的构建起到至关重要的作用[25]。
Otsuki等[26]最早将碳、氮稳定同位素技术应用于生态学中。近年来,在淡水湖泊、江河、海洋中相继出现食物网构建及营养级确定的相关报道[28-29]。食物网中的生物营养级(Trophic Level,TL)计算公式[29]如下所示:
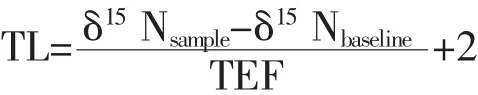
式中,δ15Nsample为系统消费者氮稳定同位素比值;δ15Nbaseline则为该系统基准生物的氮稳定同位素比值;TEF为营养级的富集度(Trophic Enrichment Factor);2是基线生物的营养级。
基线生物一般选择营养级较低的初级消费者,而选取的不同将导致营养级产生差异,常见的有浮游动物[30]、贻贝[31]、毛蚶[32]、缢蛏[33]、栉孔扇贝[22]等;不同学者也使用各基线生物营养级的均值来计算[35-36],但这些动物的数据单一且变动太大,导致结果无法得到精确。此外,不同海域所对应的基线生物不同,亦或是海域食物网的构建需要以多个基线生物为基础,目前尚无统一结论。因此,基线生物的确定仍需要标准化和规范化。
动物组织中的δ15N容易在体内富集,营养级越高,其δ15N值也越大。Peterson[10]通过室内模拟生物生长试验得到营养级富集度为3.4‰;蔡德陵等[31]通过人工培养鳗鱼试验得到营养级富集度为2.5‰;闫松光等[32]在2014年取二者均值2.95‰,并于2016年由Qu等进行了检验和校正,仍确定为2.5‰[23]。这3个值计算营养级的结果存在差异,与海域环境、食性来源和生物体长密切相关[36]。营养级富集度的确定是一个长期的过程,毕竟不同生物体内的营养级富集度可能有所差异,在今后还需要加进试验方面的研究。
迄今为止,碳、氮稳定同位素技术已经频繁出现于国内外海洋食物网基础和营养级的报道中。Du等[37]沿用质量平衡建模和稳定同位素技术构建了亚热带海湾的营养结构,发现使用质量平衡建模得出的结果数值偏低;Bin等[38]使用碳、氮稳定同位素技术分析并讨论了佐格湿地夏季水生生物群落食物网结构特征,得出该群落内各个生物的δ13C和δ15N数值存在较大差异。营养级的差异性与生物体长规格密切相关,种内差异现象甚至超过种间差异[39],使得构建食物网基础和确定生物营养级仍存在较大外延拓展的研究空间。
2 稳定同位素技术在食性来源方面的应用
稳定同位素方法是根据消费者稳定同位素比值与其食物同位素比值相接近的原则来判断生物的食物和有机质来源[40],确定不同的动物类群、消化率和水文条件,进而确定食物贡献。多数海洋生物在个体发育阶段存在一次甚至多次的食性转变现象[35],Willis等[41]运用碳、氮稳定同位素技术探究了缅因州近岸海湾鱼类种群可能的食物来源,发现食性转变现象不仅与生物所处环境相关,还与生物不同发育时期、个体大小有关[42]。因此,早期有学者提出食源贡献比例的方式来计算生物的食物来源[43],Phillips等[16]使用贝叶斯模型(Baysian isotope mixing model)加强改进了IsoSource软件,为同位素数据的处理和分析提供了有效手段。该软件运用n种同位素确定n+1种食物来源,计算公式如下:

式中,δ13Nconsumer为消费者的某一种稳定同位素比值,δ13NA、δ13NB和 δ13NC分别为生物 A、B 和 C 的某一种同位素比值,FA、FB和FC分别为3种可能的食物来源A、B、C的贡献比。
近年来,国内外碳、氮稳定同位素技术被广泛运用在淡水、海域等不同水体的生态系统之中。Chen等[44]利用稳定同位素技术构建了澳门湿地的食物网基础,发现该地鱼、虾、蟹物种的食性来源主要包括有机碎屑、底栖动物和腐殖质等;Lamontagne等[45]利用贝叶斯模型研究了半干涸河口3种主要鱼类的食性来源,阐明在河流有机物质输入缺乏的情况下,通过海洋内源输入也有助于维持河口生态系统;Willis等[41]探究了缅因州近岸海湾鱼类种群可能的食物为浮游生物、底栖动物、甲壳类及小型鱼类等,为该海域食物分布的时空结构变化提供了基础数据和理论参考。
3 稳定同位素技术在营养群落结构方面的应用
海洋食物网与水环境间的物质循环及能量流动,海洋生物能够相互依存、相互制约[22,29]。通过海洋生物的营养群落结构、时空分布及群落多样性等,为海域生态环境及生态系统评价提供了基础资料[46]。目前,稳定同位素技术的成熟使其被广泛应用于生态系统食物网构建及营养关系动态变化等研究领域[49-50],对全面开展海洋生物食物网的研究工作及了解海洋生物群落结构具有重要意义。
稳定同位素技术在量化指标上的应用成为一种趋势。量化指标不仅能预测中小范围内的系统结构变化[49],还能定量分析海洋生态系统的营养群落结构变化[29]。Layman等[50]在2007年提出了δ13C-δ15N的二维坐标,计算了6个营养结构的量化指标,以坐标系中的散点代表群落内各物种的生态位。“δ空间”被定义为同位素生态位,用以划分生物在营养级上的细小差别[50]。
目前,有不少国内外学者对不同海域的生物群落结构、群落多样性做了研究和报道。纪炜炜等[51]利用稳定同位素生态位的方法分析东海北部海域内主要游泳动物营养结构的季节性变化,证明该海域生物群落的食源多样化与营养冗余程度密不可分;Beld等[47]使用定量稳定同位素方法研究了维多利亚湖食物网结构的异质性,发现杂食性鱼类以无脊椎动物为食源,群落的营养结构与季节变化、空间分布密切相关;Preciado等[48]使用胃含物分析和稳定同位素方法分析了深海海底生物群落的食物网功能及特点,发现游泳生物比深海生物更易受群落功能的影响,为营养生态及海洋食物网构架提供了基础理论和科学依据。
4 结论与展望
从众多研究者近年来取得的一系列重要成果中,碳、氮稳定同位素技术主要应用于海洋生态系统食物网构建基础及其动态变化,确定海洋生物营养级连续值及追溯其食性来源,以及评估海域生态环境评价指标和分析生物营养群落结构变化,还包括基线生物的选取和营养级富集度的确定。
但目前由于海洋环境及生态系统遭受不同程度的破坏,海洋食物链不断缩短、营养回路减少,极大降低了食物网的稳定性[52]。因此,更多碳、氮稳定同位素技术研究将侧重于水域环境因子与生物间的营养关系以及食物网结构的时空变化[53]。并且,未来的研究方向也可能结合多种分析方法与计算机算法,利用同位素质量守恒模型[16]和食物网模型[37],以“简化食物网”的目的为前提来确定优势物种或关键种的营养级,以此构建海域食物网基础及动态关系。这种方法将越来越广泛运用于海洋生态系统的研究中。
猜你喜欢
当代水产(2022年8期)2022-09-20
昆明医科大学学报(2022年2期)2022-03-29
当代水产(2021年10期)2022-01-12
中南林业科技大学学报(2021年11期)2021-12-17
食品安全导刊(2021年20期)2021-08-30
分析化学(2017年5期)2017-06-21
教育教学论坛(2016年49期)2017-02-27
河北渔业(2014年11期)2014-11-17
河北渔业(2014年8期)2014-08-27
同位素(2014年3期)2014-06-13
