
一株欧洲类禽H1N1猪流感病毒的分离鉴定与反向遗传系统的建立
2019-07-06 06:11胡晓鲁兰海玲李雪松杨健美滕巧泱陈鸿军李泽君刘芹防关平原
中国动物传染病学报 2019年3期
胡晓鲁,张 婷,兰海玲,3,李雪松,杨健美,滕巧泱,陈鸿军,李泽君,刘芹防,关平原
(1. 内蒙古农业大学兽医学院,呼和浩特010018;2. 中国农业科学院上海兽医研究所,上海200241;3.黑龙江大学生命科学学院,哈尔滨150080)
猪流感(swine influenza,SI)是由猪流感病毒(swine influenza virus,SIV)引起的一种严重的呼吸道传染病,其特点是急性、热性和高度接触性,临床以高热、呼吸困难、咳嗽、衰竭、迅速康复或死亡为特征[1]。猪流感病毒属正黏病毒科、A型流感病毒属,为单股、负链、分节段RNA病毒,基因组由8个独立RNA片段构成[2]:PB1、PB2、PA、NP、HA、NA、M和NS,分别编码聚合酶(PB1、PB2和PA)、核蛋白(NP)、血凝素(HA)、神经氨酸酶(NA)、基质蛋白(M1和M2)、核输出蛋白(NEP)以及非结构蛋白(NS1和NS2)[3-4]。
1930年,美国首次报道了猪流感,现已遍布世界各地[5]。目前,全球流行的猪流感病毒主要亚型为H1N1、H1N2和H3N2 。H1N1亚型病毒分为古典型和类禽源型,H3N2亚型病毒为类人源型,H1N2亚型病毒由H1N1、H3N2亚型病毒重组产生[6-7]。1930年首次分离到猪流感病毒属经典H1N1亚型[8],1979年分离到欧洲类禽H1N1猪流感病毒[9]。1969年中国台湾省首次分离到猪流感病毒,1970年病毒传到香港和大陆。由于猪呼吸道上皮细胞具有唾液酸α2,3 受体和α2,6 受体,所以猪被认为是人对禽流感病毒适应过程的中间宿主,同时也是流感病毒重组和复制的“混合器”[10]。猪流感病毒不经过重组就能感染人和禽类,在对流感病毒流行、跨种传播的研究以及公共卫生方面,猪流感病毒都具有重要意义[11],成为流感病毒研究的热点[12]。8质粒拯救系统是利用RNA 聚合酶Ⅰ和RNA 聚合酶Ⅱ启动转录的双向系统,其特点是完全以质粒为基础,不需要任何辅助质粒,避免了大量的筛选工作,对病毒基因组的操作更加方便,成为拯救流感病毒的重要工具,大大促进了对病毒致病机理的研究与疫苗研发[13]。
山东省某发病猪场猪群有明显的高热、咳嗽、呼吸困难等症状,发病率较高,死亡率低。本研究采集了猪咽喉棉拭子,并成功分离到1株H1N1亚型猪流感病毒,序列分析结果显示此毒株为欧洲类禽猪流感H1N1亚型病毒。体外复制试验结果显示,分离病毒在ST细胞上可以高效繁殖,而在MDCK细胞中复制水平较差。利用RT-PCR将分离病毒的8个基因片段扩增后克隆到8质粒拯救系统中,并成功拯救出该株猪流感病毒,测序结果显示拯救病毒与野生毒株序列完全一致。反向遗传系统的建立为欧洲类禽H1N1亚型猪流感病毒基因组结构与功能之间的关系,流感病毒跨种传播的机制以及新型猪流感疫苗株的进一步研究奠定基础。
1 材料与方法
1.1 细胞与质粒293T细胞、MDCK细胞、ST细胞、pLLB-A、pLLB-G双向表达流感病毒反向遗传系统载体为本实验室保存。9~11日龄SPF鸡胚购自北京维通利华实验动物技术有限公司。
1.2 主要试剂反转录试剂购自TaKaRa公司;PCR试剂(Phanta)和ClonExpress® II重组克隆试剂盒购自南京诺唯赞生物科技有限公司;限制性内切酶STUI、DNP1购自NEB公司;大肠杆菌DH5α感受态细胞购自北京全式金生物技术有限公司;胎牛血清(FBS)、DMEM培养液、MEM培养液、Opti-MEM培养液、青霉素链霉素混合液(SP)和TPCK胰酶购自美国Gibco公司;牛血清白蛋白(BSA)购自Sigma公司;转染试剂Mirus购自BIO公司;RNA提取试剂盒购自Tiangen公司;凝胶回收试剂盒和小剂量质粒抽提试剂盒购自Axygen公司;大剂量质粒抽提试剂盒购自Qiagen公司。
1.3 病毒分离鉴定采集发病猪咽喉棉拭子,加入2 mLPBS,离心收集上清。取100 μL上清(其余保存于-80℃冰箱)经尿囊腔接种9~11日龄SPF鸡胚,37℃孵育,收集48 h后未死亡有血凝活性的鸡胚尿囊液,保存于-80℃冰箱备用。
1.4 反向遗传质粒的构建使用RNA提取试剂盒提取病毒RNA,利用反转录试剂盒将其反转录为cDNA,以通用引物进行PCR扩增目的基因,电泳后胶回收扩增产物。将PLLB-A、PLLB-G载体经StuI内切酶酶切,电泳后进行胶回收。利用重组克隆试剂盒,将目的片段与载体的胶回收产物进行同源重组,转化大肠杆菌DH5α感受态细胞,12 h后挑取单个菌落进行菌液PCR鉴定。阳性质粒送公司测序,选取无突变的克隆进行质粒大量提取,测定浓度后用于病毒拯救。

表1 PCR引物信息Table 1 Sequence of PCR primers in this study
1.5 病毒全基因组测序与Blast分析将构建好的反向遗传质粒,使用载体引物pLLB-F、pLLB-R测序。利用SeqMan软件组装基因组,对全基因组进行Blast,分析与分离病毒最接近的病毒亚型与来源。
1.6 病毒拯救使用含有10%FBS和1%SP的DMEM培养液培养293T细胞,将细胞传至6孔板。当细胞密度超过80%时,进行转染。将8 μg质粒混合物(1 μg/质粒)和24 μL转染试剂分别溶于250 μL OPTIMEM无血清培养基中,室温静置5 min后相互混匀,结合30 min。弃掉6孔板中的细胞生长液,PBS轻洗1次, 再加入1.5 mL OPTI-MEM无血清培养基,将质粒与转染试剂的混合物 (共500 μL)均匀加入,在37℃、5%CO2的细胞培养箱内培养48 h。转染的同时,设立只加2 mL OPTI-MEM无血清培养基作为对照组。
转染48 h后,在96孔血凝板中测定血凝滴度。取50 μL细胞上清与50 μL PBS混匀,进行8个滴度的倍比稀释后,每孔加入50 μL 0.7 %鸡红细胞,静置30 min后观察结果。
1.7 拯救病毒在MDCK与ST细胞中复制能力比较用含有10%FBS和1% SP的DMEM培养液培养ST细胞,以含有5% FBS和1% SP的MEM培养液培养MDCK细胞,将细胞传至6孔板。当细胞密度超过80%时,进行接毒。弃掉6孔板中的细胞生长液后用PBS轻洗细胞2次,每孔加入1.5 mL维持液(MDCK为4%BSA、1% SP、1‰ TPCK胰酶的MEM培养液;ST为4%BSA、1% SP、1‰ TPCK胰酶的DMEM培养液);然后每孔再加入500 μL转染后的细胞上清,在37℃、5% CO2的细胞培养箱内培养48 h,连续传3代。接毒48 h后,收集细胞上清测定血凝滴度。若有血凝活性,将病毒收集,冻存于-80℃冰箱。
为了比较分离病毒在不同细胞中的复制能力,将MDCK细胞、ST细胞分别传至T25细胞瓶。当细胞密度超过80%时,每瓶用PBS洗2次后加入维持液5 mL。按照接毒剂量稀释病毒至1 mL,加入瓶中,置于37℃、5%CO2的细胞培养箱内培养。分别在第12、24、36、48 h,每瓶取细胞上清300 μL保存至-80℃,然后每瓶补充300 μL细胞维持液,每个时间点做3个重复。最后对每个时间段收取的样品测定TCID50,利用GraghPad Prism 5 Project软件画图。
2 结果
2.1 病毒分离鉴定将处理后的样品接种9~11日龄SPF鸡胚,72 h后收集鸡胚尿囊液。检测结果显示病毒的HA滴度为22,具有血凝活性。将HA阳性的尿囊液接种MDCK细胞,接种后72 h,出现细胞病变,细胞变圆、脱落,而对照组细胞状态保持完好(图1)。MDCK细胞上清进行血凝活性测定,结果为22。
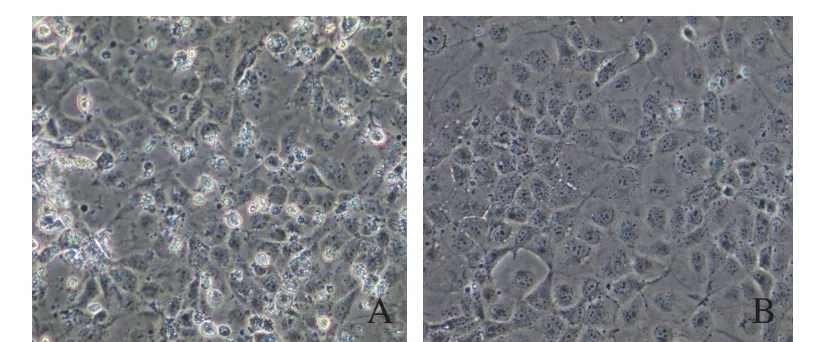
图1 分离到猪流感病毒在MDCK细胞中产生细胞病变Fig.1 Cytopathic effect(CPE) caused by SIV in MDCK cells
2.2 分离病毒的基因扩增与质粒构建结果使用流感病毒通用引物对病毒cDNA进行PCR,可扩增出8个目的基因片段(图2~5)。将反向遗传质粒pLLB载体进行酶切。原质粒为超螺旋结构,因此扩增条带在线性化质粒的下方,电泳结果显示酶切成功(图6)。回收酶切后的PLLB载体,分别与扩增的8个基因片段进行连接,转化大肠杆菌DH52感受态细胞,然后进行菌液PCR鉴定。鉴定结果为阳性的菌液送公司测序,并对测序结果进行Blast比对。结果显示8个片段均来源于欧亚类禽猪流感分支,表明质粒构建成功。从阴性菌液中提取质粒,测定浓度和OD值后分装,进行病毒拯救。

图2 分离毒株NP、PA基因片段PCR 扩增结果Fig.2 PCR result of NP and PA gene segments from isolated strain
图3 分离毒株PB1、PB2基因片段PCR 扩增结果Fig.3 PCR result of PB1 and PB2 gene segments from isolated strain

图4 分离毒株HA、NA基因片段PCR 扩增结果Fig.4 PCR result of HA and NA gene segments from isolated strain
2.3 分离病毒全基因组测序与Blast分析Blast结果显示这8个基因片段与欧洲类禽猪流感H1N1病毒的相似性均大于等于95%(表2)。其中PB1、HA、M、NS基因都与A/Jiangsu/ALS1/2011(H1N1)较为接近,说明这两个毒株存在较近的亲缘关系。
2.4 病毒拯救结果及验证将拯救的病毒接种在MDCK细胞上,48 h后出现细胞病变,传3代后,上清中血凝效价为22。将拯救病毒接种到ST细胞上,48 h后出现细胞病变,传3代后,上清中的血凝效价为26,说明本分离毒株更适应猪的ST细胞。将拯救病毒进行全基因组测序,结果显示拯救的病毒与野生型的病毒序列完全一致,将拯救的毒株命名为A/swine/Shandong/1/2016(H1N1)。

图5 分离毒株M、NS基因片段PCR 扩增结果Fig.5 PCR result of M and NS gene segments from isolated strain

图6 反向遗传质粒PLLB载体酶切鉴定结果Fig.6 Identification of endonuclease reaction
表2 分离毒株8个基因片段的同源性分析Table.2 Homology analysis of genes fragments from isolated strain by Blast
2.5 病毒在MDCK与ST细胞中复制能力比较为了进一步比较分离毒株在不同细胞中的复制能力,将拯救的病毒分别接种到MDCK和ST细胞,不同时间点收集样品进行滴度测定。结果显示本研究中分离的病毒在ST细胞中复制水平显著高于在MDCK细胞中复制水平,感染MDCK后12 、24、36 h,上清中的病毒滴度均低于TCID50检测下限(TCID50)。生长曲线结果显示本分离毒株可以很好的适应ST细胞,但在MDCK细胞中适应性较差,具体的分子机制需要进一步研究。
图7 分离毒株在不同细胞上的生长曲线Fig.7 Growth dynamics of rescues virus in ST cells and MDCK cells
3 讨论
猪流感具有世界性发生和流行性的特点,遍及世界各大洲。由于猪对禽流感和人流感病毒都易感,不同病毒同时感染猪后可在猪体内发生重排,生成具有新基因型的病毒,因而猪被认为是新型流感病毒的混合器。2009年发生的甲型H1N1流感病毒也是猪-人-禽流感病毒的三源重配病毒。因此,进行猪流感的监测和病毒特性的系统研究对流感大流行的预警具有重要的意义。
2000年以前,我国大陆猪群中较少有猪流感暴发的新闻报道。2000年以后,猪流感的活动性明显增强,某些地区的抗体阳性率高达80%。中国猪群中主要流行的猪流感病毒包括经典H1N1亚型、欧洲类禽H1N1亚型、H1N2亚型以及H3N2亚型流感病毒。1991年我国猪群中首次分离到经典H1N1亚型SIV[14],2000年后经典H1N1 SIV的流行性显著提高,但2007年后欧洲类禽H1N1病毒逐渐取代经典H1N1病毒成为我国猪群中的优势毒株[15-16]。调查结果显示,2009年大流行的甲型H1N1、经典H1N1、欧洲类禽H1N1以及北美三源重排H3N2等多种亚型SIV在我国猪群中共流行,不同病毒之间的基因重排现象频繁发生。2009大流行的甲型H1N1流感病毒爆发后迅速回传到猪群,并与猪群中流行的猪流感病毒发生基因重排[17-19]。我国多个地区的猪群中均分离到欧洲类禽H1N1 SIV与H1N1/2009的重排病毒。研究表明,欧洲类禽猪流感H1N1病毒不仅能在猪之间有效传播,而且可以通过气溶胶方式在雪貂间传播,其潜在的分子机制尚不清楚[20]。
MDCK细胞是流感病毒敏感细胞系,但本研究中分离的毒株在MDCK细胞中的复制能力较弱,具体原因与机制需要进一步研究。本研究成功构建的分离毒株的反向遗传学系统,为研究欧洲类禽H1N1猪流感病毒的复制及致病性分子机理提供了工具。
猜你喜欢
动物医学进展(2022年9期)2022-11-26
中国典型病例大全(2022年11期)2022-05-13
医学概论(2022年3期)2022-04-24
临床医学工程(2022年3期)2022-04-20
首都食品与医药(2022年3期)2022-02-21
科学大观园(2022年2期)2022-01-23
文萃报·周二版(2021年47期)2021-12-14
中国现代医药杂志(2019年9期)2019-10-31
祝您健康·文摘版(2019年2期)2019-06-11
科学24小时(2019年5期)2019-06-11
