
黄粉甲储存蛋白hexamerin基因的克隆及表达分析
2019-07-03 09:17:24徐志文任雪敏刘乃勇吴国星朱家颖
西南林业大学学报 2019年4期
徐志文 任雪敏 赵 满 刘乃勇 吴国星 杨 斌 朱家颖
( 1. 西南林业大学云南省森林灾害预警与控制重点实验室,云南 昆明 650233;2. 云南农业大学植物保护学院,云南 昆明 650201)
储存蛋白(SP)为昆虫生长发育提供重要的营养,该蛋白首次于1969年从处于变态阶段的丽蝇(Calliphora erythrocephala)体内被提取并鉴定,命名为丽蝇蛋白(Calliiphorin)[1]。因其是由6个相同或者相似的亚基聚合而成的球形六聚体,通常被称作hexamerin(六聚体蛋白)。Hexamerin是昆虫体内普遍存在的一种特异性血淋巴蛋白,其分子量在60~110 kDa之间,大多在80 kDa左右[2-5]。它与节肢动物酪氨酸酶、甲壳纲的血蓝蛋白、双翅目芳基蛋白受体具有相同的起源,属于血蓝蛋白超家族,被认为是由血蓝蛋白的含铜载氧体失去载氧功能进化而来[4,6-7]。它通常在幼虫或若虫的脂肪体中合成并释放到血淋巴中,在老熟幼虫的血淋巴中浓度及含量达到最大,待幼虫化蛹时又以蛋白颗粒的形式重新被脂肪体选择性吸收进入脂肪体[8-9]。
储存蛋白hexamerin是蛋白质和氨基酸的储存库[1,8]。并且,hexamerin在昆虫生长发育、免疫和雌性卵发育等过程中发挥着重要作用,还能直接参与表皮的形成、滞育、亲脂性物质运输、能量代谢以及变态[10-13]。如在红头丽蝇(C. uicina)、家蝇(Musca domestica)、烟草天蛾(Manducasexta)等昆虫中,hexamerin参与几丁质表皮组织的构建和蛋白质组成[11,14];在马铃薯甲虫(Leptinotarsa decemlineata)[15]和始红蝽(Pyrrhocoris apterus)[16]中,hexamerin与成虫滞育有关。此外,研究还发现hexamerin参与亲脂性物质运输[17],特别是参与疏水的脂溶性杀虫剂运输[18],部分hexamerin还参与能量代谢[2]。虽然hexamerin有诸多功能,但是在完全变态昆虫中,其最主要的功能是作为储存蛋白为幼虫变态发育提供营养物质[19],为成虫生长发育提供氨基酸和能量[8]。
黄粉甲(Tenebrio molitor)作为饲养管氏肿腿蜂(Sclerodermus guani)的重要替代寄主,为人工繁蜂及释放管氏肿腿蜂防治林木蛀干害虫提供了重要的价值。同时,黄粉虫也是重要的经济昆虫和仓储害虫,而且能危害天然纤维,如蔺草(Schoenoplectus trigueter)、芦苇(Phragmites communis)等编织物,对我国蔺草制品出口产业造成巨大威胁。因其具有容易饲养、适合遗传杂交、易开展RNAi实验等优点,所以可作为理想的模式昆虫。目前,就黄粉甲贮藏蛋白的研究而言,仅在其幼虫中发现了1个分子量为86 kDa的包囊蛋白与在滞育中起重要作用的储存蛋白具有很高的相似性[20]。为此,本研究以黄粉甲为试验材料,克隆获得了2个hexamerin基因(命名为TmolHex1和TmolHex2),并对其序列和表达特征进行了分析,研究结果为今后揭示hexamerin的生物学功能提供参考。
1 材料与方法
1.1 供试昆虫
供试昆虫饲养参照Zhu等[21]的方法,黄粉甲为在实验室以饲料饲养繁殖多代的种群。
1.2 基因克隆
利用TRIzol(Invitrogen)试剂,参照试剂盒说明书提取黄粉甲蛹总RNA。总RNA经分光光度法测定其含量和纯度后,采用1%琼脂糖凝胶电泳检测总RNA的完整性。以提取的高质量RNA为模板,用SMARTTMRACE cDNA amplification kit (Clontech)合成cDNA模板。以实验室前期对黄粉甲cDNA文库测序获得的hexamerin基因3′端序列为依据,用Primer Premier 5.0分别设计TmolHex1的 5′ RACE(5′-CCGTAGTACTGTTGTTTGTACCTGT-3′)和TmolHex2的 5′ RACE(5′-CGTTGGCATAGTAGAACAACTT-3′)特异性引物。参照RACE试剂盒说明书,PCR扩增获得基因的5′端序列。PCR反应条件为:94 ℃预变性 3 min;94 ℃ 变性 35 s,60 ℃ 退火 30 s,72 ℃延伸2 min 30 s,40个循环;72 ℃延伸10 min。将获得的PCR产物用1%琼脂糖凝胶电泳检测,切取的目的条带纯化后送至上海杰李生物技术有限公司测序。
1.3 序列分析
将实验室前期对cDNA文库测序得到的序列和克隆测序获得的序列,拼接得到TmolHex1和TmolHex2的基因全长。利用在线软件ORF Finder(www.ncbi.nlm.nih.Gov/gorf/gorf.html)预测基因的开放阅读框,DNAMAN预测分子量及等电点,Genetyx软件将其基因的核苷酸序列翻译成氨基酸序列并分析其组成成分,SignalP 4.1 Server (http://www.cbs.dtu.dk/services/SignalP/)进行信号肽预测,NetNGlyc 1.0 Server分析氨基酸序列N-糖基化位点,ClustalX 1.83软件进行多序列比对。
1.4 荧光定量PCR
选择刚蛹化1 d的黄粉甲蛹,将蛹放置于冰上取其表皮、脂肪体和血细胞3种组织,同时收集黄粉甲低龄幼虫(1~3龄幼虫混合样品)、高龄幼虫(7~9龄幼虫混合样品)、老熟幼虫(12~14龄幼虫混合样品)、蛹化1 d的蛹和羽化1 d的成虫。每组样品收取3个生物学重复,收取的材料放置于盛有TRIzol试剂的1.5 mL离心管中,研磨后保存在-80 ℃超低温冰箱中备用。
采用TRIzol(Invitrogen)试剂提取黄粉甲不同发育阶段和蛹不同组织样品的总RNA,提取的总RNA去除DNA后,使用RevertAid First Strand cDNA Synthesis Kit(Thermo)反转录合成cDNA模板。根据克隆获得的TmolHex1和TmolHex2序列,设计TmolHex1的荧光定量正向引物(5′-AGTCATCCAGCAAGCATACAG-3′) 和 反 向引 物(5 ′-TAGAAGGAGTTGAGGCGTA-3′) 及TmolHex2的正向引物(5′-ATTACGCACAGAAGCAGAAAG-3′)和反向引物(5′-CAAATGGTAGTTGTGGGAGAT-3′),以 18S RNA基因(正向引物 5′-TTTCAAATGTCTGCCTTATC-3′和反向引物 5′-TGTGGTAGCCGTTTCTCA-3′)作为内参,使用Rotor Gene-Q荧光定量PCR仪检测2个基因在黄粉甲不同发育阶段和蛹期不同组织的表达水平,每个处理重复3次。PCR反应条件为:95 ℃预变性2 min;95 ℃变性15 s,55 ℃退火30 s,45个循环。荧光定量PCR数据采用2-ΔΔCT方法进行计算[22],同一处理下的基因相对表达量用SPSS软件进行单因素方差(One-way ANOVA)统计分析。
2 结果与分析
2.1 基因克隆及序列分析
克隆获得的TmolHex1(GeneBank登录号:KF734080) 和TmolHex2( GeneBank登 录 号 :KF734081)的开放阅读框(ORF)的长度分别为2 133 bp和2 103 bp,可编码氨基酸分别为710个和700个,5′端非编码区分别为12 bp和28 bp,3′端非编码区分别为125 bp和109 bp。其推导的氨基酸序列预测理论分子量分别为85.53 kDa和84.35 kDa,理论等电点分别为6.05和6.72。TmolHex1和TmolHex2均存在 2个N-糖基化位点。SignalP分析发现,TmolHex1和TmolHex2的氨基酸序列中均存在信号肽序列,分别为MKLLVAILALGAFACTYPQVQI和MRLILVALLAGLCAVSA。对 TmolHex1和 TmolHex2的氨基酸组成进行分析发现(表1),TmolHex1蛋白中的甲硫氨酸和芳香族氨基酸含量分别为0.14%和22.25%(苯丙氨酸5.63%、酪氨酸16.20%、色氨酸0.42%),而TmolHex2蛋白中的甲硫氨酸和芳香族氨基酸含量分别为2.57%和21.86%(苯丙氨酸6.86%、酪氨酸13.86%、色氨酸1.14%),表明它们属于芳香族储存蛋白。

表 1 TmolHex1和TmolHex2氨基酸组成成分分析Table 1 Analysis of amino acid composition of TmolHex1 and TmolHex2
多序列比对分析发现,TmolHex1与赤拟谷盗(Tribolium castaneum)、柞蚕(Antheraea pernyi)、黑带二尾舟蛾(Cerura vinula)的hexamerin氨基酸一致性分别为58%、32%、32%和32%(图1),而TmolHex2与这些昆虫hexamerin氨基酸一致性分别为72%、31%、31%和32%(图1)。Tmol-Hex1和TmolHex2具有保守的芳香贮存蛋白家族特征序列,其中TmolHex1的为YFTEDVRLNSFYYYYNIYYP和 TSLRDPAFY,TmolHex2的为YYMEDVGLNSFYYYYNLYYP和TSLRDPAFY。
2.2 表达特征
在不同发育阶段,TmolHex1基因在高龄幼虫和老熟幼虫中的表达量明显高于其他发育阶段的表达量,蛹中的表达量次之,低龄幼虫和成虫期的表达量最低(图2)。TmolHex2基因在老熟幼虫和蛹期的表达量明显高于其他发育阶段的表达量,高龄幼虫的表达量次之,低龄幼虫和成虫期的表达量最低(图2)。在蛹期不同组织中,TmolHex1和TmolHex2基因在脂肪体中的表达量最高,血细胞中的表达量次之,而表皮中的表达量最低(图3)。
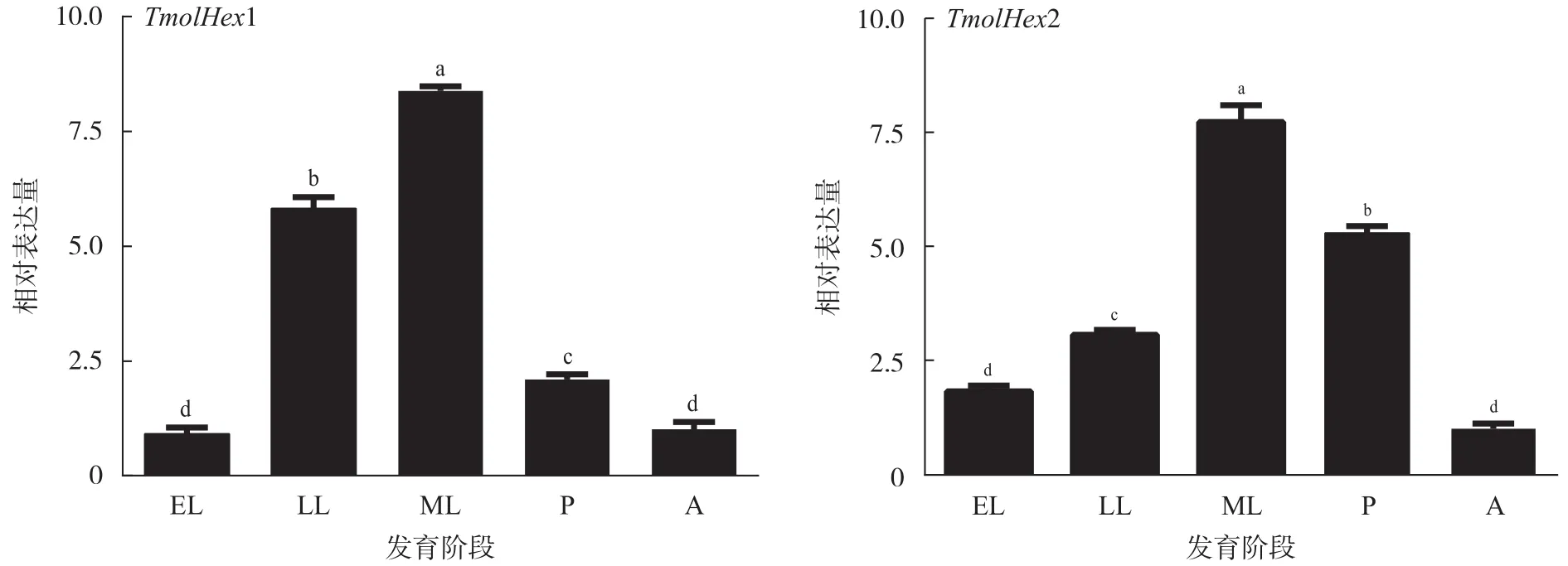
图 2 TmolHex1和TmolHex2基因在不同发育阶段的相对表达量Fig. 2 Relative expression levels of TmolHex1 and TmolHex2 genes of T. molitor at different developmental stages

图 3 TmolHex1和TmolHex2基因在蛹期不同组织中的相对表达量Fig. 3 Relative expression levels of TmolHex1 and TmolHex2 genes in different tissues of T. molitor pupae
3 结论与讨论
在现今的研究报道中,研究者以hexamerin中芳香族氨基酸、甲硫氨酸、酪氨酸等重要氨基酸的含量为依据,可将其划分为4类:1)芳香蛋白,酪氨酸和苯丙氨酸的总含量为16%~25%,而甲硫氨酸的含量少于3%[23];2)富甲硫氨酸蛋白,甲硫氨酸的含量为4%~11%,而芳基氨基酸含量小于15%[24];3)双翅目幼虫血清蛋白(Larval serum protein 2, LSP-2),芳香氨基酸和甲硫氨酸的含量比丽蝇蛋白(calliphorin)中的含量少[25];4)其他,如高甲硫氨酸和酪氨酸含量的芳基蛋白、核黄素结合蛋白和保幼激素抑制蛋白等[2,26]。 基 于 hexamerin 的 分 类 , TmolHex1 和TmolHex2富含芳香族氨基酸,且氨基酸序列中存在芳香家族贮存蛋白的保守序列[12,27],表明TmolHex1和TmolHex2隶属于芳香族类。这一结果与目前报道的鞘翅目hexamerin均为芳香族氨基酸的结果相符[28-29]。
TmolHex1和TmolHex2基因在老熟幼虫和蛹期的脂肪体中表达量高,该研究结果与在桔小实蝇(Bactrocera dorsalis)、米蛾(Corcyra cephalonica)、大豆食心虫(Leguminivora glycinivorella)、家蝇等昆虫中的报道相似[30-32]。同时,研究还发现TmolHex1和TmolHex2基因在黄粉甲蛹期和成虫期的表达量相对于老熟幼虫中的表达量显著较低,且在成虫期的表达量相对于蛹期也明显下降。现有研究认为,贮存蛋白一般由取食的幼虫合成,在蛹期以蛋白质晶体储存在脂肪体细胞中,到成虫期几乎全部降解为氨基酸,从而认为它的功能可能为蛋白质和氨基酸的储存库,并参与成虫蛋白质的合成,被降解得到的肽类或氨基酸可为成虫发育和生殖所利用[2,5,33-35]。TmolHex1和TmolHex2基因在不同发育阶段和蛹组织中的表达特征表明,它们可通过高表达得以在老熟幼虫的血淋巴中大量积累,并可在幼虫化蛹过程中又以蛋白颗粒的形式重新被脂肪体选择性的吸收进入脂肪体[8-9]。为此,推测认为,它们的主要功能应该是作为储存蛋白为昆虫提供所需的氨基酸和能量,参与黄粉甲的变态发育。and Physiology, 1990, 15(1): 33-41.
猜你喜欢
食品科学(2023年13期)2023-08-12 00:41:48
中国生物化学与分子生物学报(2022年7期)2022-09-07 05:28:48
航天制造技术(2022年3期)2022-07-15 06:04:02
中国药科大学学报(2021年6期)2021-12-31 03:05:20
环境昆虫学报(2020年3期)2020-07-07 01:35:48
果农之友(2019年12期)2019-02-06 04:03:33
现代农村科技(2018年8期)2018-08-23 12:07:50
金色少年(奇趣科普)(2017年6期)2017-07-25 09:21:14
分析测试学报(2015年9期)2015-12-17 16:44:28
湖南农业科学(2015年5期)2015-02-27 14:33:51
