
陆地棉转录因子Gh WRKY70的克隆及表达分析
2019-06-11 06:13刘冬梅娄喜艳裴冬丽
西南农业学报 2019年5期
刘冬梅,娄喜艳,喻 拥,李 瑶,裴冬丽*
(1. 商丘师范学院生物与食品学院植物与微生物互作省级重点实验室,河南 商丘 476000;2. 商丘工学院土木工程系,河南 商丘 476000)
【研究意义】棉花是重要的经济作物。 陆地棉是世界上种植最广泛的棉花种类,占全棉产量的90 %。 异源四倍体物种衍生自作为父本Gossypiumraimondii,提供了D基因,以及作为母本的A基因组物种G.Arboreum。他们在五千至一千万年前分化,在一百至两百万年前重新团聚起来形成陆地棉。
【前人研究进展】转录控制是细胞或生物体调节其基因表达的主要机制,转录因子(TFs)是通过结合其启动子区域来调节几个其他基因的表达,从而导致转录的蛋白质。除了正常的基因转录控制之外,它们还可以作为对环境信号的响应来调节基因表达[1]。WRKY转录因子是目前发现的最大的转录因子之一,有研究表明,WRKY蛋白在防御包括细菌,真菌和病毒病原体在内的各种生物胁迫的植物防御中发挥关键作用,它们在发育过程中也起着重要的调控作用,如毛状体启动,胚胎形态发生,衰老以及植物激素如赤霉酸,脱落酸,或水杨酸。也有越来越多的证据表明WRKY蛋白参与了对各种非生物胁迫的反应。在拟南芥中,微阵列分析显示,一些WRKY转录物在响应各种非生物胁迫如盐度,干旱和寒冷时受到强烈调控。在水稻中,非生物胁迫(寒冷,干旱和盐度)或各种植物激素处理下,54个WRKY基因在转录本丰度上表现出显着差异。在大麦中,WRKY基因Hv-WRKY38在受到寒冷和干旱胁迫时表达,而在大豆中至少有9种WRKY基因在非生物胁迫下差异表达[2],值得注意的是,单个WRKY基因可以通过与几种VQ蛋白(具有VQ连接的基序,包括SIB1和SIB2(西格玛因子相互作用蛋白))调节多种非生物胁迫来调节多种非生物反应[3]。
自从第1个编码WRKY蛋白SPF1的cDNA 由 Ishiguro 和 Nakamura在甘薯中克隆出来后,更多的WRKY基因在不同植物中被发现,尤其是在禾本科植物中,包括野生燕麦,水稻,小麦等[4]。近年来,随着高通量测序工作的实施,许多物种的测序工作都已经完成,棉花WRKY基因家族包含116个基因[5]。WRKY蛋白因含有约60个氨基酸的1个或2个域指定的WRKY结构域而得名。在其N端含有一段保守的氨基酸基序WRKYGQK,在C端含有一个新型的C2H2(C-X4-5-C-X22-23-H-X-H,X可以是任意的非保守氨基酸)或C2HC(C-X7-CX23-H-X-C)锌指结构[6]。根据其结构域的数量和锌指结构的类型,WRKY蛋白可分为3类。I类有2个WRKY结构域和C2H2型锌指结构域,II类有1个WRKY结构域和C2H2型锌指结构域,根据其结构可以进一步分为5个亚类(II-a、II-b、II-c、II-d和II-e),III类有1个WRKY结构域和 C2HC型锌指结构[7]。WRKY结构域对于的不同顺式作用DNA元件显示出高结合亲和力称为W盒(T/CTGACC/T)[8]。W盒已经在许多涉及防御和应对环境压力的基因的启动子中鉴定出,W盒也存在于WRKY基因启动子本身中,表明WRKY基因是自我调节的,并且还可以调节应激相关基因的表达[9]。
有研究表明,WRKY70积极参与油菜素类固醇(BR)调节的生长,并且负面地参与干旱反应[10]。鹰嘴豆CaWRKY70通过调控CaHDZ12表达,从而影响植株的抗旱和耐盐能力[11]。以拟南芥为模型系统,WRKY70通过激活SA信号通路调控由蜡状芽孢杆菌AR156引发的诱导型全身抗性(ISR)[12]。油菜BnWRKY70基因表达程度与其植株的抗病性呈正相关[13]。拟南芥中的WRKY70同时参与了防御丁香假单胞菌侵染和RPP4调控的抗霜霉病过程,并通过与RCY1抗病蛋白作用来调节拟南芥抗黄瓜花叶病毒[14]。陆地棉WRKY3在根、茎和叶中形成并通过调节各种植物激素(包括水杨酸、脱落酸、茉莉酮酸甲酯、乙烯、赤霉素等)进行表达[15]。对WRKY转录因子在二倍体A和D棉花品种中的综合比较研究中, 总共鉴定了112G.raimondii和109G.arboreumWRKY基因。在2种物种之间没有发现显着的基因结构或结构域改变,但许多SNP在外显子和内含子区域分布不均匀[16]。据已有文献显示,WRKY转录因子还参与棉花花药的发育[17]。在野生大豆中。WRKY基因家族成员GsERKY20参与调控植物开花时间[18]。NAC和WRKY的共同表达在调控芝麻MS中起关键作用[19]。在拟南芥中,WRKY34转录因子已被证明参与其花粉的发育[20],表明该转录因子在不同植物体中的功能基本相似,推测GhWRKY70也有上述部分或全部功能。
【本研究切入点】根据前人研究结果,WKY70转录因子参与多个生物学过程,其中包括调控开花时间。转录组测序结果显示,WKY70转录因子在陆地棉洞A在不育系不育花药中表达高于可育花药,推测该基因与棉花雄性不育相关。本文克隆了该基因并对其结构和亲缘关系进行了分析,同时通过实验验证该基因在雄性不育和可育花药中的差异表达,为了进一步研究该基因功能奠定基础。
【拟解决的关键问题】该基因的克隆和结构分析为棉花雄性不育基因功能和雄性不育机理的研究提供前提,也为棉花雄性不育材料的创造提供基础。
1 材料与方法
1.1 实验材料
1.1.1 植物材料 陆地棉洞A不育系花药(花蕾d=3~4 mm)。
1.1.2 试剂 DL2000 DNA Marker购自全式金公司, ExTaq聚合酶、rTaq聚合酶、solution1 连接酶、pMD18-T载体,M购自TaKaRa公司,DNA 凝胶回收试剂盒购自东盛公司,Trizol试剂购自上海闪晶分子生物科技有限公司,ReverTra Ace qPCR RT Kit试剂购自东洋纺(上海)生物科技有限公司,大肠杆菌由本实验室保存。
1.2 方法
1.2.1 RNA提取与cDNA的合成 RNA提取参照Trizol(上海闪晶)试剂说明书操作步骤。
cDNA合成参照ReverTra Ace qPCR RT Kit(东洋纺)试剂说明书操作步骤。
1.2.2GhWRKY70转录因子的克隆 利用Primer 5设计GhWRKY70转录因子全长引物,引物序列Unigene19749-F:NNNNGGATCCATGGGAAGTGTGTCAGCTTGGC;Unigene19749-R:NNNNG TCGACCTA GACCAACTCACTTTCATCAAACTG N(A,T,C或G)。
GhWRKY70转录因子全场基因PCR扩增体系为:10×ExTaqbuffer 5 μl;10 mM dNTPs 1 μl;Primer-s 1 μl;Primer-as 1 μl;cDNA 1 μl;0.5 μl;加ddH2O至 50 μl。其PCR扩增程序为:预变性94 ℃,5 min;变性94 ℃,30 s;退火63 ℃,1 min;延伸72 ℃,50 s;终延伸72 ℃,10 min;进行35个循环,4 ℃保存。
1.2.3 PCR产物T载体的连接 将克隆所得片段使用试剂盒进行胶回收,并与pMD18-T连接。T载体连接体系solution1 5 μl;片段 4 μl;载体1 μl,20 ℃过夜连接。
1.2.4 大肠杆菌感受态细胞的制备及载体转化 方法参照张丽霞,贾海燕(2013)一种简便高效大肠杆菌感受态细胞制备方法[21]。
1.2.5 转化后大肠杆菌的克隆及测序 以菌液为模版进行克隆,用Primer 5程序设计菌液PCR检测引物H-19749-1-F GGTTGGGATTTTGGGGTTGC;H-19749-1-R GACTGGAAGATGCTGGCTCC。菌液检测PCR体系10×rTaqbuffer 2 μl;10 mM dNTPs 0.2 μl;Primer-s 0.3 μl;Primer-as 0.3 μl;菌液 1 μl;rTaq0.2μl;ddH2O 16 μl。其PCR扩增程序为:预变性94 ℃,5 min;变性94 ℃,30 s;退火60 ℃,40 s;延伸72 ℃,50 s;终延伸72 ℃,10 min;进行35个循环,4 ℃保存。将结果送上海生物工程公司测序,通过多次比对序列,最后得出完整序列。
1.2.6 电泳 ① 琼脂糖凝胶制备:称取0.4 g的琼脂糖,加入40 mL的TBE,放入微波炉中加热溶解,煮沸3次至完全溶解,待胶凉至60~70 ℃时,倒平板凝固。② 点样:将RNA提取液(PCR扩增产物或待检测的菌液)和Loding Buffer按照5∶1的比例混匀点入点样孔,点2个样。③ 电泳条件:电压120 V,电泳15 min。④ 染色:跑完电泳后,胶片在EB中浸泡30 min。⑤ 观察:紫外光下观察电泳条带。
1.2.7 预测与分析 通过NIBI在线软件ORF Finder(https://www.ncbi.nlm.nih.gov/orffinder/)预测GhWRKY70氨基酸个数以及蛋白质序列。用在线软件SMAR(http://smart.embl-heidelberg.de/)对蛋白质结构域分析。
利用Expasy数据库中的在线软件Prot Param(http://web.expasy.org/protparam/)分析并预测GhWRKY70蛋白的理化性质。
用swiss-model分析软件(http://swissmodel.expasy.org//SWISS-MODEL.html)进行三级结构预测。
1.2.8 引物设计 通过Primer 5设计引物,转录因子GhWRKY 70在不育和可育花药中的表达进行Q-PCR检测,以棉花泛素蛋白基因为内参用,差异表达分析引物如下:H-1975-F:GCCTCCAATTGTCCCCTTCA;H-1975-R:TACGCAAACTGTTCACCCGA,片段长度161 bp。根据导出数据计算各指标的平均Ct值、ΔCT值及ΔΔCT值,并计算检测指标的相对表达水平。
2 结果与分析
2.1 RNA提取及反转录
从图1可以看出,条带清晰,没有拖带现象,RNA无降解,分光光度计测定RNA浓度为175.08 ng/μl,纯度(A260/A280)为2.147,可用于基因克隆模板。
2.2 Gh WRKY70全长基因的克隆
从图2可以看出,其位置处于750~1000 bp中间,片段大小与目的基因相符,条带清晰。
2.3 检测菌液PCR
根据菌液PCR检测结果(图3),挑取阳性克隆,送公司测序。

M:DL2000 Marker;1:棉花不育花药总RNA图1 RNA提取及反转录Fig.1 RNA extraction and reverse transcription
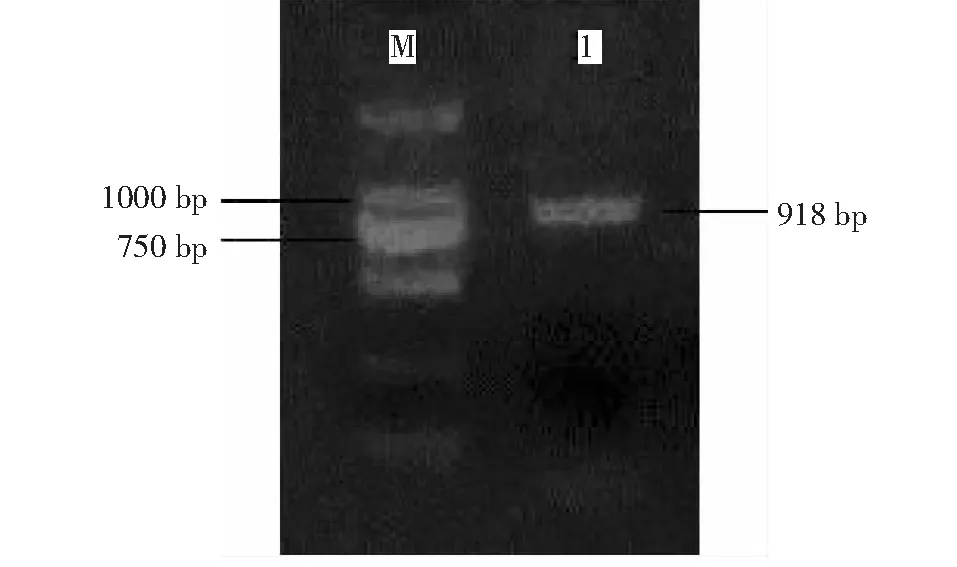
M:DL2000 Marker ;1:PCR所得基因克隆产物图2 Gh WRKY70基因全长的克隆Fig.2 Full-length gene cloning of Gh WRKY70
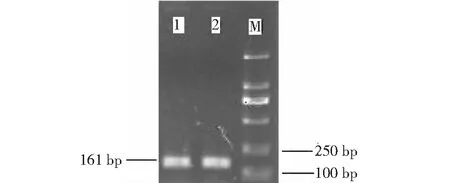
M:DL2000 Marker;1~2:PCR所得菌液克隆产物图3 检测菌液PCR Fig.3 Detection of bacteria PCR
2.4 测序及提交
将获得的全长基因送上海生物工程公司测序,得到GhWRKY70完整编码区序列,该基因全长为918 bp,将测序所得到的GhWRKY70完整编码区序列通过BackIt提交到NCBI,Gene BANK号为MF278614。
2.5 蛋白质序列结构预测
2.5.1 Gh WRKY70蛋白质序列GhWRKY70基因编码305个氨基酸,该蛋白只有1个WRKY结构域,包含61个 aa其位置处于128~191,E值为4.44e-36,在292~301处还有1个组成低复杂性序列区域(图4)。
2.5.2 Gh WRKY70蛋白质的二级结构预测 Gh WRKY70蛋白二级结构预测结果显示该蛋白质α螺旋(Hh)为98个,占全部结构的32.13 %,延伸链(Ee)37个,占全部结构的12.13 %,13个β转角(Tt),占全部结构的4.26 %,无规则卷曲有157个(Cc),占全部结构的51.48 %(图5)。
2.5.3 Gh WRKY70蛋白质三级结构预测 在对Gh WRKY70进行蛋白质三维结构预测的结果中显示,该蛋白只有1个WRKY结构域,与蛋白质结构域分析的结果一致,说明预测结果准确。将生成的 PDB 结果在 SWISS-pdb Viewer 软件中观察蛋白质的结构,结果表明Gh WRKY70三维空间结构主要是由β-折叠和无规则卷曲组成(图6)。

图4 蛋白质结构域分析Fig.4 Protein domain analysis
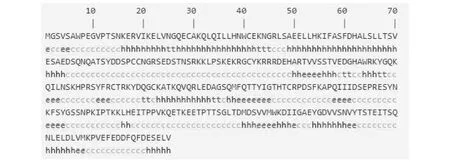
图5 GhWRKY70蛋白的二级结构预测Fig.5 Secondary structure prediction of GhWRKY70 protein
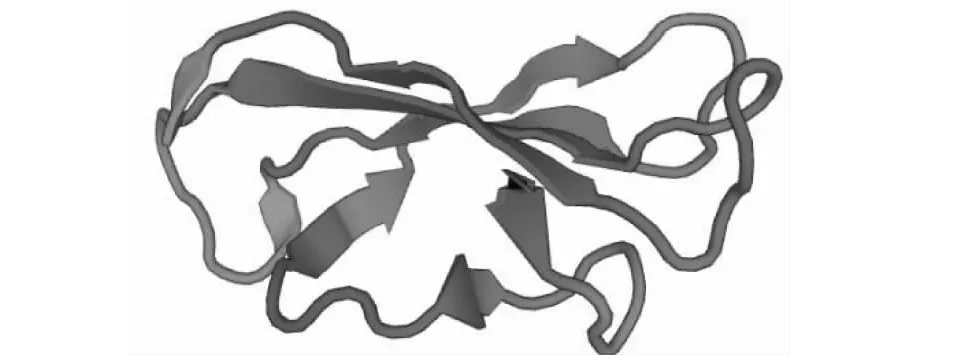
图6 Gh WRKY70蛋白质三级结构Fig.6 Gh WRKY70 protein tertiary structure
2.6 蛋白质理化性质
在Gh WRKY70蛋白理化性质预测分析结果中显示,该蛋白有305个氨基酸,由19种氨基酸组成,分子量:34648.65,理论等电点:5.97,分子式:C1502H2366N426O490S13。在WRKY70的氨基酸序列中Ser的使用频率最高,为10.5 %。WRKY70蛋白总的负电荷残基数为46个,正电荷残基数为41个,蛋白质的不稳定指数较高,为50.81,指数大于40,表明该蛋白是不稳定的。WRKY70蛋白亲水性较强,氨基酸残基疏水性总和(GRAVY)为-0.838。
2.7 同源性分析及系统进化树的构建
运用MEGA5软件对陆地棉(MF278614GossypiumhirsutumWRKY70)和烟草(KM510344.1NicotianatabacumWRKY70)、拟南芥(AF421157.1ArabidopsisthalianaWRKY70)、小麦(LC068769.1TriticumaestivumWRKY70)、甘蓝(KM593152.1Brassicaoleraceavar WRKY70)、芸苔(KF430076.1BrassicarapaWRKY70)、辣椒(KF484401.1CapsicumannuumWRKY70)、玉米(NM_001154348.1ZeamaysWRKY70)、水稻(BK005073.1OryzasativaWRKY70)、大豆(NM_001315509.1GlycinemaxWRKY70)、粗山羊草亚种(XM_020312323.1Aegilopstauschiisubsp WRKY70)、芭蕉亚种(XM_009417198.2Musaacuminatasubsp WRKY70)、可可(XM_018129667.1TheobromacacaoWRKY70)、亚洲棉(XM_017749682.1GossypiumarboreumWRKY70)、西瓜(GQ453670.1CitrulluslanatusWRKY70)15个序列进行同源性分析, 使用邻近法(NJ method), bootstrap 设为 1000,构建进化树,其结果显示GhWRKY70与亚洲棉的系统进化树在同一支上,同源性最高,可可在另一个分支上,相比之下同源性次之。水稻和玉米在最远的分支上,则与其同源性最差(图7)。
2.8 Gh WRKY70基因在不育和可育花药中的差异表达分析
从图8可以看出,GhWRKY70基因在不育花药中的表达显著高于可育花药,表明该基因可能与洞A雄性不育有关,Q-pCR结果与转录组测序的结果一致,验证了转录组测序结果的重要性。

图7 陆地棉Gh WRKY70基因的同源进化树分析Fig.7 Analysis of Gh WKY70 gene in upland cotton based on homologue evolutionary tree analysis
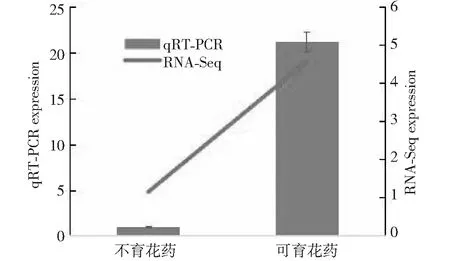
图8 GhWRKY 70基因在不育和可育花药中的差异表达Fig.8 Differential expression of GhWRKY 70 in sterile and fertile anther
3 讨 论
通过本次研究,对GhWRKY70基因有了一个初步的了解。该基因全长918 bp,根据软件分析,预测编码305个氨基酸。在GhWRKY70编码的蛋白中,只有1个WRKY结构域和1个C2-HC(C-X7-C-X23-H-X1-C)型的锌指结构,属于第III类,该类转录因子只在高等植物中存在[22]。开花植物中的WRKY III基因被认为起源于单子叶植物和真核生物的分歧。拟南芥III组成员的时间表达分析支持这些成员是不同植物防御信号传导途径的一部分,包括相容,不相容和非宿主相互作用,表明它们的功能分化。因此,WRKY III基因在植物适应和进化中发挥了关键作用。 WRKY III基因被认为是进化方面最先进的,并且在适应性方面最为成功的[23]。该蛋白WRKY结构域中高度保守的WRKYGQK这七个氨基酸序列并不是一成不变的,有些植物的WRKY成员的这七个序列中的W、Q和K会出现变异,而且Q位点突变频率最高[24]。拟南芥WRKY 结构域中存在的Q突变为K。水稻WRKY结构域中存在Q突变为E。WRKY结构域的突变会改变蛋白质的结构,从而导致蛋白质的某些功能的减弱甚至丧失。下一步工作将基因转入陆地棉或拟南芥中,验证该基因功能,为创造棉花雄性不育材料奠定基础。
猜你喜欢
麦类作物学报(2022年8期)2022-08-08
湖北农业科学(2022年11期)2022-07-18
河北农业大学学报(2022年2期)2022-04-26
园艺与种苗(2020年12期)2021-01-08
实用肿瘤学杂志(2020年4期)2020-12-08
蔬菜(2019年5期)2019-05-21
江苏农业科学(2016年8期)2017-02-15
飞碟探索(2015年9期)2015-11-05
红领巾·探索(2015年9期)2015-09-10
天津农业科学(2015年9期)2015-09-02
