
水稻骨干恢复系川恢907抗稻瘟病基因聚合与精准改良
2019-06-11 06:22白玉路张志勇徐登武蒲志刚王闵霞
西南农业学报 2019年5期
白玉路, 王 平,张 琼,张志勇,徐登武,蒲志刚,王闵霞
(四川省农业科学院生物技术核技术研究所, 四川 成都 610066)
【研究意义】稻瘟病是水稻的主要病害之一,每年都会造成大量的损失。生产实践表明,利用抗稻瘟病亲本培育并种植抗稻瘟病品种是控制稻瘟病最经济有效的方法[1]。然而水稻稻瘟病菌生理小种变异快,使得具有单一抗病基因的品种容易丧失抗性,长期实践证明,利用优良的抗性基因资源,进行基因聚合拓宽其抗谱是培育水稻抗病亲本与品种的关键,也是对抗水稻病害最为经济、安全和有效的方法。【前人研究进展】目前,至少有69个抗稻瘟病位点共84个主效基因已被定位,这些基因成簇地分布于除第3染色体外的所有水稻染色体上。前人研究表明,Pi2、Pita以及Pik是目前重要的稻瘟病抗性基因位点,其在抗稻瘟病育种材料选育方面具有重要应用价值,育种家已经利用Pi2、Pik以及Pita等基因成功选育出一系列抗稻瘟水稻新材料[2-12]。近年来,多个研究人员将Pi1、Pi2、Pigm、Pi40、Pita、Pib、Pi9以及Pi49等抗稻瘟病病基因与其它抗病、米质相关基因进行聚合,创制出了一系列高产、优质、多抗水稻新材料,为培育优质、多抗水稻新品种奠定了丰富的材料基础[13-17]。【本研究切入点】川恢907是四川省农业科学院生物技术核技术研究所通过远缘杂交、辐射诱变结合花药培养技术选育出的大穗高配合力强优势水稻三系骨干恢复系,含有抗稻瘟病基因Pik和广谱抗病基因Pita,已配出多个杂交水稻新品种通过国家和省级审定,具有良好的应用价值[18]。多年来,川恢907与田间稻瘟病菌生理小种的不断互作,其抗性逐年下降,配制的杂交水稻组合稻瘟病抗性愈来愈差,限制了其在生产上的推广应用。为了保持川恢907的持久抗病性,本研究以含有广谱抗稻瘟病基因Pi2的优质恢复系五山丝苗为抗稻瘟病基因供体,川恢907为拟改良受体,通过常规杂交、回交和自交,以及基于高分辨率熔解曲线(HRM)的分子标记辅助选择技术(MAS),将Pi2基因导入到骨干恢复系川恢907中,聚合抗稻瘟病基因Pik、Pita和Pi2。【拟解决的关键问题】通过本研究实现抗病基因有效聚合累加,拓宽抗谱提高其抗稻瘟病能力,并保持其他农艺性状不变,实现川恢907稻瘟病抗性的精准改良,延续其强优势配合力,为选育出更多的优良杂交水稻新品种提供广谱抗病的高配合力恢复系。
1 材料与方法
1.1 供试材料
Pi2基因受体亲本及轮回亲本川恢907,含有抗稻瘟病基因Pik、Pita,由四川省农业科学院生物技术核技术研究所自育;Pi2基因供体亲本五山丝苗,由广东省农业科学院水稻研究所提供;稻瘟病感病对照CO39,由四川省农业科学院植物保护研究所提供。
1.2 抗稻瘟病基因分子检测
基于HRM技术体系的Pik、Pita和Pi2基因分子标记参考罗文龙等开发的功能型分子标记[19](表1),并采用巢式PCR 进行扩增。每个位点分别设计外部和内部引物进行2 轮PCR。巢式PCR 的第1 轮PCR 扩增,以基因组DNA 为模板,利用外部引物扩增;扩增体系:2×TaqMaster Mix 5 μl,External Primer 0.3 μl,模板DNA0.6 μl,用ddH2O补足10 μl体系;PCR反应程序为95 ℃ 3 min;然后35个循环:95 ℃ 15 s,55 ℃ 30 s,72 ℃ 1 min; 最后72 ℃延伸7 min,4 ℃保存。待反应完成后,向PCR 管分别加入50 μl双蒸水,以稀释PCR产物(约5倍)。第2 轮PCR,扩增体系:2×TaqMaster Mix 4 μl,Internal Primer 0.25 μl,模板DNA 0.6 μl,荧光染料Evagreen 0.4 μl,高温、低温内标各1 μl,用ddH2O补足10 μl体系;PCR反应程序为95 ℃ 3 min;然后20个循环:95 ℃ 15 s,57 ℃ 25 s,72 ℃ 10 s; 最后95 ℃解链2 min,40 ℃ 1 s。
1.3 抗稻瘟病基因聚合改良系的培育
以五山丝苗作为Pi2基因供体,与被改良材料川恢907杂交,再以川恢907为轮回亲本,连续回交5代获得BC5F1,再自交3代获得BC5F4。从BC1F1开始,每代连续采用基于HRM体系的Pik、Pita和Pi2基因功能型分子标记进行辅助选择,筛选出含有Pik、Pita和Pi2基因且与川恢907农艺性状相似的单株材料进行连续回交、自交,直到完全稳定。
1.4 抗稻瘟病基因聚合改良系DNA指纹图谱鉴定
根据农业行业标准NY/T 1433-2014中48对SSR引物对基因聚合改良系和川恢907进行指纹图谱鉴定。通过随机选取混合样,采用CTAB方法提取基因组DNA,PCR反应体系为10 μl,10×Taqbuffer 1.0 μl (Mg2+),2 mmol/L dNTPs 1.0 μl,10 μmol/L正反向SSR引物各0.75 μl,模板DNA 20 ng。反应程序为:95 ℃预变性5 min;94 ℃DNA变性1 min,55 ℃退火30 s,72 ℃延伸30 s,35个循环;72 ℃终延伸7 min。扩增产物在8 %非变性聚丙烯酰胺凝胶中恒压电泳,银染检测[20]。

表1 抗稻瘟病基因分子标记引物序列
1.5 抗稻瘟病基因聚合改良系稻瘟病田间抗性鉴定
2017-2018年将从BC5F4筛选获得的聚合了3个抗病基因Pik、Pita和Pi2并且DNA指纹图谱一致的的改良系、川恢907及稻瘟病感病对照CO39在邛崃天台山镇稻瘟病抗性鉴定圃进行稻瘟病田间抗性鉴定,4月中旬播种,5月中下旬移栽,每个材料设3次重复,每次重复30穴,采用稻瘟病自然诱发接种鉴定,全生长期不施用杀菌剂,肥水管理按一般高产栽培管理。于稻瘟病盛发期调查各材料叶瘟及穗颈瘟发病率、损失率、综合抗病指数。以发病最为严重的重复作为该材料的最终调查数据。
1.6 抗稻瘟病基因聚合改良系主要农艺性状差异分析
2018 年4月,将改良系与川恢907种植于四川省农业科学院现代农业科技创新示范园,试验实行随机区组排列,设置 3 次重复,每个重复100穴。栽插密度为行距30 cm、株距17 cm,常规田间管理。每个重复定点取3个单株,调查生育期、株叶型、穗型、粒型和株叶颜色等农艺性状,对株高、剑叶长、剑叶宽、穗长、穗粒数、实粒数、结实率、千粒重等主要农艺性状进行差异性比较,并通过T检验方法分析改良系与川恢907之间主要农艺性状差异显著性。
2 结果与分析
2.1 抗稻瘟病基因聚合改良系的培育与抗病基因检测
2014年夏季在四川省农业科学院现代农业科技创新示范园配制川恢907×五山丝苗杂交F1组合,2014年秋季种植于四川省农业科学院英州南繁基地,抽穗前通过HRM分子检测将含Pik、Pita和Pi23个目的单株,以川恢907作轮回亲本配制川恢907//907/五山丝苗回交组合,接着2014年冬季在英州南繁基地种植回交组合,2015年春季抽穗前通过HRM分子检测全部单株,选择含Pik、Pita和Pi23个目的基因且外观农艺性状与川恢907基本相似的部分单株再与川恢907进行回交,每年每个世代如此操作。每年成都夏季种植1季,海南秋天和冬天分别种植1季,每季种植回交组合10个以上,单株在200株以上,直到2015年秋季在海南种植BC4F1,在种植的株系中选择出3个群体基本稳定、农艺性状较一致的株系进行抗病基因检测,选取部分单株初步进行DNA相似性分析,共检测出5个单株与川恢907的差异位点为0或1个,2015年冬季在海南选择含有3个目的基因并且无差异位点的单株再与907回交获得BC5F1,接着对回交的组合在成都和海南连续自交3代,自交每代进行相似性鉴定,筛选无差异位点的进行自交,直到2017年春季获得BC5F4, 经DNA相似性检测和农艺性状鉴定,从中筛选获得1份含有3个纯合目的基因且外观农艺性状与川恢907一致且农艺性状稳定的抗稻瘟病基因聚合改良系69235-4-10,定名为新川恢907。
2.2 新川恢907 DNA指纹图谱鉴定
以农业行业标准NY/T 1433-2014中48对SSR引物对25株新川恢907与原始被改良材料川恢907进行DNA指纹图谱鉴定。结果表明,2个材料48对引物的扩增产物带型表现一致,新川恢907与川恢907之间DNA指纹图谱无差异。部分分子标记对BC5F4材料与川恢907的相似性鉴定见图1。
2.3 新川恢907稻瘟病田间抗性鉴定
2017-2018年,分别对川恢907、新川恢907以及感病对照CO39在邛崃市天台山镇稻瘟病抗性鉴定圃进行田间抗性鉴定。通过对叶瘟发病率、叶瘟病级、穗颈瘟发病率以及损失率的调查,结果表明:新川恢907叶瘟发病率、叶瘟病级、穗颈瘟发病率、损失率以及综合抗病指数显著低于被改良材料川恢907(表2)。由此可以看出,多基因聚合显著提高了川恢907的田间稻瘟病抗性。

1~2:2株川恢907单株;3~27:25株新川恢907单株1-2: Two plants of Chuanhui 907; 3-27:Twenty-five plants of new Chuanhui 907图1 部分分子标记对新川恢907与川恢907 DNA指纹图谱鉴定 Fig.1 Identification of partial SSR molecular markers between new Chuanhui 907 and Chuanhui 907
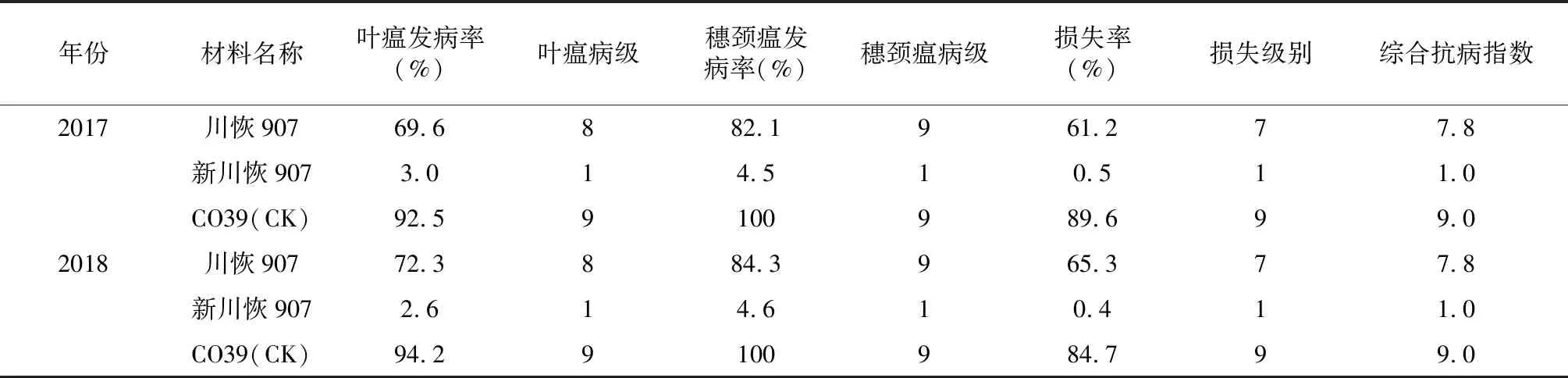
表2 新川恢907田间抗性鉴定结果
2.4 新川恢907与川恢907之间主要农艺性状差异分析
通过对新川恢907与川恢907的农艺性状调查,新川恢907全生育期150.6 d,川恢907全生育期150.2 d,二者相当;其株叶型、叶色等外观农艺性状也无明显差异;同时,对株高、穗长、穗粒数、实粒数等12个农艺性状拷种测量并进行T检验,比较其差异显著性,结果表明:在12个测量的主要农艺性状中P值在0.1~0.5之间的性状有8个,大于0.5的性状有4个。新川恢907与川恢907之间各外观农艺性状不存在显著差异(图2,表3)。
3 讨 论
通过利用携带不同抗稻瘟病基因的材料,采用杂交、回交和分子辅助选择技术结合聚合抗病基因改良水稻稻瘟病的抗性,培育出广谱抗稻瘟病的材料是可行的。黄艳玲等[21]、向聪等[22]、裘烨等[23]通过分子辅助选择技术分别将Pi9基因、Pigm基因、Pi-kh基因导入感病的两系不育系或恢复系中,选育出了稻瘟病抗性显著增强的不育系或恢复系材料。本研究采用杂交、回交、自交与基于HRM的分子标记辅助选择技术相结合的方法,将抗稻瘟病基因Pi2导入到含有抗稻瘟病基因Pik和Pita基因的川恢907中,选育出了稻瘟病抗性显著提高的新川恢907。
通过杂交、多次回交、自交,结合分子标记辅助选择技术和基因型相似性分析,能精确改良优良水稻恢复系的抗性,并能保持原材料的全部优良性状。在以往的报道中,基本上都是通过杂交或回交,结合分子辅助选择技术,选育出抗病性增强的新的育种材料,对骨干恢复系或不育系的精准改良罕见报道。川恢907精准改良的成功为其他恢复系抗性改良乃至不育系抗性、米质等精准改良上提供了可行的借鉴,其新川恢907和原川恢907所配组合的杂种优势及抗性表现等研究工作正在进行中。

图2 新川恢907与川恢907对比Fig.2 Comparion between new Chuanhui 907 and Chuanhui 907
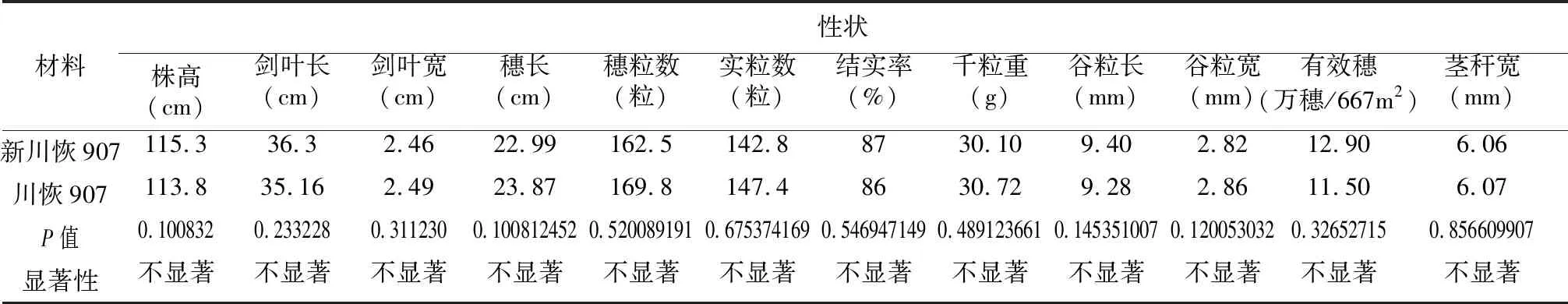
表3 新川恢907与川恢907主要农艺性状差异显著性分析
对优良水稻品种和材料的精确改良,需要不断发掘新的优良基因,开发新的分子标记和检测技术。为了保持优良水稻品种持久抗病性,就需要对新发现的抗稻瘟病基因,通过杂交、回交和分子辅助选择技术不断进行基因聚合累加、结合基因型相似性分析做出精确改良。在改良川恢907中,回交了5次,自交了3次,如何加快材料的改良速度,提高选育效率,回交和自交多少代次才能保证和原材料基因型相似,将进一步进行研究。
4 结 论
本研究以携带有含广谱抗病基因Pi2的五山丝苗为供体,以携带有抗稻瘟病基因Pita、Pik的川恢907为受体,通过上述方法对川恢907的稻瘟病抗性进行了精确改良,获得了含有3个目的基因的改良恢复系新川恢907,以NY/T 1433-2014中48对SSR引物对其进行指纹图谱鉴定,背景与川恢907无差异,通过T检验主要农艺性状无明显差异,并且稻瘟病抗性较川恢907显著提升,表明川恢907精准改良获得成功。
猜你喜欢
智慧农业导刊(2022年21期)2022-11-03
当代水产(2022年7期)2022-09-20
河北科技师范学院学报(2022年2期)2022-08-26
作物学报(2022年6期)2022-04-08
中国饲料(2021年17期)2021-11-02
空间科学学报(2021年1期)2021-05-22
落叶果树(2021年6期)2021-02-12
甘肃教育(2020年18期)2020-10-28
天津农林科技(2020年3期)2020-08-13
四川蚕业(2020年2期)2020-07-10
